تهیه کتابخانه ژنی
تکنولوژی DNA نوترکیب
از سال ۱۹۷۰ میلادی به بعد پیشرفت های قابل توجه ای در روش ها و تکنیک های مورد استفاده برای مطالعه فرایندهای بیولوژیکی در سطح مولکولی حاصل شد. این روش ها و تکنیک ها اکثراً منجر به ایجاد ابزارهای جدید و قدرتمندی برای جداسازی آنالیز و دستکاری اسیدهای نوکلئیک شده و در حقیقت با اطلاعات فراهم شده تحول شگرفی در نواحی مختلف علوم بیولوژی همانند ،بیوتکنولوژی کشف داروها پزشکی مولکولی و ژن درمانی به وجود آورد.
کشف اندونوکلئازهای محدود کننده در اوایل ۱۹۷۰ میلادی یک کشف کلیدی بود که نه تنها امکان آنالیز مؤثر DNA بلکه قابلیت برش مولکول های DNA مختلف را فراهم ساخت به طوری که بتوان آنها را مجدداً به یکدیگر متصل و قطعات DNA نوترکیب جدید را به وجود آورد. به این فرآیند امروزه اصطلاحاً کلون سازی ژن گفته می شود . فرآیند کلون سازی ژنی قادر به انجام کشفیات بی شماری بوده و اطلاعات با ارزشی را از ساختمان عملکرد و تنظیم ژن فراهم میسازد. این فرآیند در ابتدا برای تهیه کتابخانه ژنی مورد استفاده قرار گرفت اما امروزه با توسعه و تصحیح به عنوان پایه اصلی اکثر آزمایشات بیوشیمی و بیولوژی مولکولی قرار گرفته است اگر چه PCR یک میانبری برای آنالیز ژن ها فراهم نموده است، اما هنوز در اکثر موارد تکنیک های کلون سازی نه تنها مفید هستند، بلکه یک ضرورت مطلق میباشند
ساخت کتابخانه های ژنی
اولین مرحله در اکثر آزمایشات کلون سازی ژن ،جداسازی و تخلیص DNA ژنومیک می باشد. هضم مولکول های DNA ژنومیک به وسیله اندونوکلئازهای محدود کننده انجام می شود این آنزیم ها با توجه به اینکه روی توالی های DNA خاصی فعالیت می نمایند، کلید کلون سازی مولکولی می باشند.
زمانی که نسخه هایی از مولکول DNA یک ارگانیسم با یک آنزیم اندونوکلئاز خاص بریده می شود.دستجات یکسانی از قطعات ایجاد خواهد شد
به طور کلی زمانی که DNA ارگانیسم های مختلف در مجاورت انزیم محدود کننده یکسان قرار میگیرند دستجات متفاوتی از قطعات به وجود خواهد آمد. به هر حال زمانی که DNA ژنومیک ارگانیسمی توسط آنزیم هضم می شود ، تعداد زیادی از قطعات کوچک ایجاد میشود که ممکن است هر یک تقریباً به اندازه یک ژن تنها باشد.
همان طور که میدانید گروهی از اندونوکلئازها، DNA را به صورت انتهای صاف (blunt) برش می زنند، در حالی که آنزیم های تحدیدی دیگر برشهای ناصافی را ایجاد کرده و نواحی تک رشته ای را در هر دو انتهای DNA برش شده ایجاد می نمایند. به این انتهاها، انتهاهای چسبنده گویند که با یکدیگر مکمل بوده و بنابراین قادرند با یکدیگر جفت شوند.
به علاوه پس از برش گروه های فسفات DNA همواره در انتهای '5 باقی میماند. در میان ۵۰۰ انزیمی که خصوصیات آنها شناخته شده است تنها بیش از ۲۰۰ جایگاه تحدیدی مختلف شناسایی شده است. انتخاب نوع آنزیم مورد استفاده به چندین فاکتور بستگی دارد.
به عنوان مثال، توالی های شناسایی ۶ جفت بازی به طور متوسط هر ۴۰۹۶ باز (6^4) اتفاق میافتند این اندازه بر اساس یک توالی تصادفی فرض شده است که در هر جایگاه آن یکی از چهار باز قرار میگیرد. بدان معنی است که اگر DNA ژنومیک با EcoRI که توالی '5GAATTC- 3' را شناسایی می نماید بریده شود احتمالاً قطعات ایجاد شده هر یک به طور متوسط 4kb طول خواهند داشت.
آنزیم هایی با توالی شناسایی هشت جفت بازی قطعات طولانی تری را تولید خواهند نمود. بنابراین ژنومهای بسیار بزرگ همانند DNA انسان معمولاً با آنزیم هایی که قطعات DNA طولانی را تولید می نمایند بریده میشوند. با توجه به اینکه در این حالت تعداد کمتری از قطعات برای کلون سازی و آنالیز بعدی ایجاد می،شود، سبب خواهد شد فرایند کلون سازی قابل کنترل تر شود.
اتصال مولکولهای DNA
قطعات DNA با انتهای چسبنده که در نتیجه برش آنزیم محدود کننده ایجاد شدهاند ممکن است با هر قطعه DNA دیگری که با همین آنزیم تحدیدی مجاور گشته و دارای انتهای یکسانی می باشند، متصل گردند.
بنابراین زمانی که این دو دسته از قطعات DNA با یکدیگر مخلوط میشوند انتهای چسبنده مکمل آنها با یکدیگر جفت شده و در نتیجه ممکن است قطعاتی با دو منشأ مختلف متصل به یکدیگر را ایجاد نمایند.
به این مولکول ها DNA نوترکیب گفته می شود . در این مسیر جفت شدن قطعات DNA با منشأ مولکولی یکسان نیز ایجاد خواهد شد که به آن اصطلاحاً اتصال مجدد گویند. به هر حال امروزه و کتورهای کلون سازی ارائه شده اند که بر این مشکل غلبه شده است.
قطعات DNA جفت شده به دلیل آنکه تنها از طریق پیوند ضعیف هیدورژنی بین تعداد کمی از بازهای انتهایی مکمل خود بهم متصل اند میتوانند جدا شوند. برای پایدار کردن این اتصال می توان DNA لیگاز در فرایندی به نام لیگاسیون استفاده نمود. معمولاً از آنزیم لیگاز جدا شده از باکتریوفاژ T4 که اصطلاحاً DNA T4 لیگاز نامیده می شود، استفاده شده که پیوند کووالانسی را بین فسفات '5 انتهای یک رشته و هیدروکسیل '3 رشته مجاور برقرار میسازد .
این واکنش وابسته به ATP بوده و اغلب در دمای 10°C انجام میگیرد تا جنبش مولکولی کمتر باشد. این دما شانس جدا شدن انتهای جفت شده قبل از اینکه توسط لیگاز پایدار شود را کاهش می دهد. به هر حال طولانی کردن زمان واکنش برای جبران فعالیت پایین DNA لیگاز در این دما لازم می باشد همچنین این آنزیم قادر است قطعات مختلف DNA با انتهاهای صاف را به یکدیگر متصل نماید.
اگر چه کارایی این واکنش بسیار کمتر از لیگاسیون انتهای چسبنده میباشد لیگاسیون مولکولهای مختلف DNA مجدداً جایگاه برش آنزیم را ایجاد می نماید به طوری که میتوان مولکول های نوترکیب تولید شده به وسیله لیگاسیون (که انتهای چسبنده داشتند) را با استفاده از همان آنزیم برش داده و قطعات اولیه مجدداً تولید گردد.
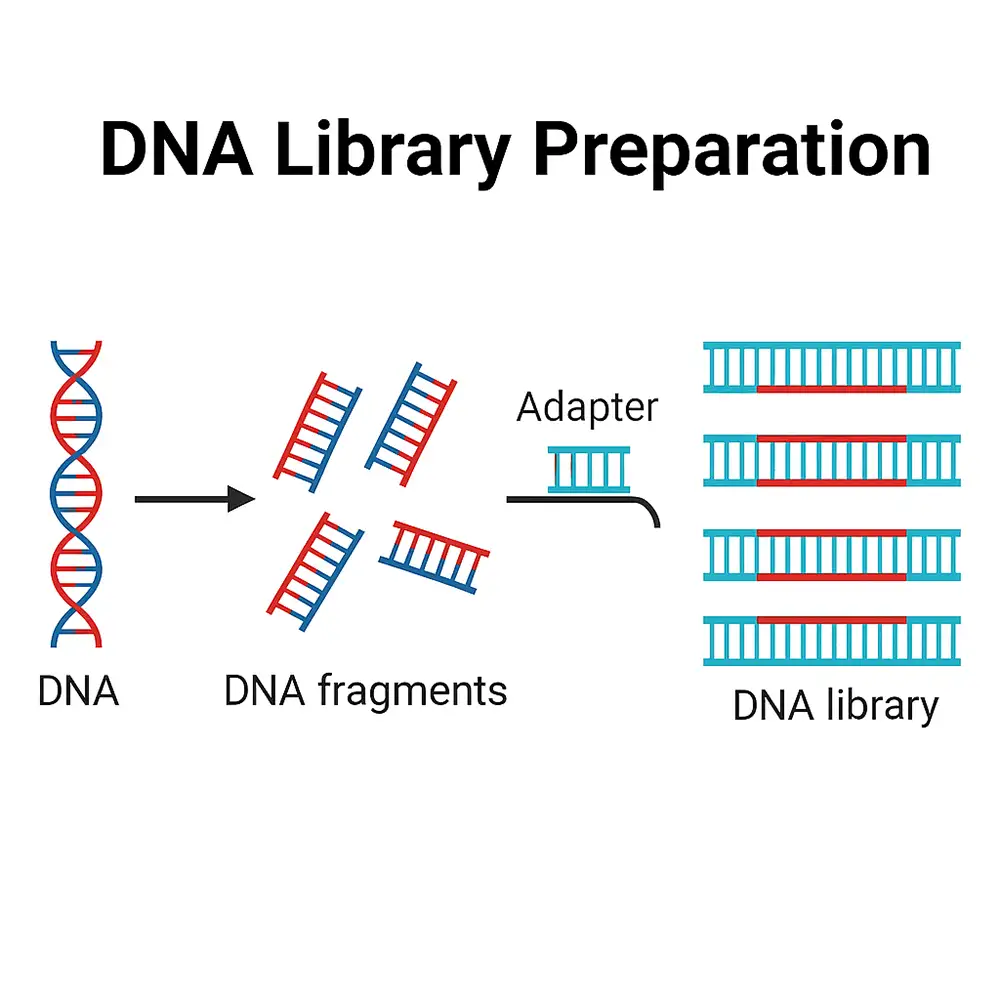
به منظور تكثير قطعه DNA هضم شده از یک ارگانیسم ضروریست آنها به یک مولکول DNA حامل خاص به نام وکتور کلون سازی متصل .گردند بنابراین لیگاسیون هر قطعه DNA با مولکول DNA وکتور اجازه میدهد كل DNA نوتركيب حاصله به طور نامحدودی در یک سلول میزبان مناسب همانند سازی نماید.
در این ،مسیر قطعه DNA مورد نظر که تکثیر شده است می تواند ماده مؤثر برای آنالیز مفصل یا دستکاری بعدی باشد. بنابراین تمامی DNA استخراج شده از یک ارگانیسم با یک آنزیم تحدیدی بریده شده و قطعات حاصله در کلکسیونی از کلون ها جای داده می شود. این کلکسیون از کلون ها، کتابخانه ژنی خوانده می.شود
نکاتی در رابطه با تهیه کتابخانه ژنی
به طور کلی دو نوع کتابخانه ژنی وجود دارد
(۱) کتابخانه ژنومیک که شامل کل DNA کروموزومال یک ارگانیسم میباشد
(۲) کتابخانه cDNA که قسمتی از mRNA یک سلول یا بافت در نقطه زمانی خاص را نشان میدهد
انتخاب نوع کتابخانه ژنی به چند چیز بستگی دارد که مهمترین آنها کاربرد نهایی قطعات DNA گرفته شده از کتابخانه می باشد. اگر هدف نهایی، چگونگی کنترل بیان پروتئین یک ژن خاص یا ساختمان آن ژن باشد، باید از کتابخانه ژنومیک استفاده نمود. امّا اگر هدف تولید پروتئین جدید یا تغییر یافته باشد یا اینکه هدف تعیین بیان اختصاصی بافت در الگوهای زمانی خاص می باشد، کتابخانه cDNA بسیار مناسب است بنابراین تفاوت اصلی در ساخت کتابخانه ژنومیک یا cDNA در ماده اسید نوکلئیک اولیه می باشد.
با توجه به اینکه ژنومیک ارگانیسم ثابت می باشد، DNA کروموزومال ممکن است تقریباً از هر نوع سلولی به منظور تهیه کتابخانه ژنومیک جدا شود، در مقابل کتابخانه های cDNA تنها mRNAهای تولید شده از یک نوع سلول خاص در زمان معینی از نمو سلولی را نشان میدهند. بنابراین توجه دقیق به نوع سلول یا بافتی که mRNA آن برای ساخت کتابخانه cDNA استفاده می شود، اهمیت دارد.
امروزه تنوعی از وکتورهای کلون سازی در دسترس می باشد که اکثر آنها از مولکولهای طبیعی همانند پلاسمیدهای باکتریایی یا ویروس های باکتریوفاژی توسعه یافته اند انتخاب وکتور نیز به نوع کتابخانه ای بستگی دارد که قرار است ساخته شود این موضوع به طور مفصل در مقالات بعدی توضیح داده خواهد شد.
کتابخانه های DNA ژنومیک
کتابخانه های ژنومیک با جداسازی کامل DNA كروموزومال از یک سلول و هضم آنها به قطعاتی با طول های دلخواه توسط اندونوکلئازهای محدود کننده ساخته می شوند
این عمل را میتوان به وسیله هضم نسبی با یک اندونوکلئاز محدود کننده که توالی چهار نوکلئوتیدی را شناسایی می نماید، انجام داد. هضم کامل با یک چنین آنزیمی تعداد زیادی از قطعات بسیار کوتاه را تولید خواهد به هر حال اگر اجازه داده شود انزیم به طور نسبی و تنها در یک تعداد کمی از جایگاههای تحدیدی قبل از توقف واکنش عمل هضم را انجام دهد هر مولکول DNA به قطعات نسبتاً بزرگی بریده خواهد شد. در چنین هضم های نسبی اندازه متوسط قطعه به غلظت نسبی آنزیم تحدیدی، شرایط و مدت اینکوباسیون بستگی دارد.
همچنین امکان تولید قطعات DNA به وسیله برش های فیزیکی نیز وجود دارد اگر چه انتهاهای قطعاتی که از این طریق حاصل می شود ممکن است نیاز به ترمیم داشته باشد تا انتهاهای صاف را ایجاد نماید این عمل به وسیله استفاده از یک DNA پلی مراز اصلاح شده به نام پلی مراز کلنو به دست میآید که در اثر هضم DNA پلی مراز I با سوبتلیسین ایجاد می شوند.
این برش یک قطعه بزرگ آنزیمی را میدهد که فاقد فعالیت '5 به '3 اگزونوکلئازی بوده اما هنوز دارای عمل '5 به '3 پلی مرازی می باشد این آنزیم شکاف های که به وسیله برش فیزیکی در هر دو انتهای '3 DNA ایجاد شده را با استفاده از dNTPهای مناسب پر می نماید.
مخلوط قطعات DNA سپس به یک وکتور متصل شده و در مرحله بعد کلون می شوند اگر کلون های کافی تولید شود احتمال بسیار بالایی وجود دارد که هر قطعه DNA خاص همانند یک (ژن) حداقل در یکی از کلونها وجود داشته باشد به طور کلی تعداد کلون ها همیشه در یک اندازه قابل کنترلی حفظ میشوند که در مورد کتابخانه های پروکاریوتی به قطعاتی 10kb نیاز می باشد. طول قطعات در کتابخانه های پستانداران باید افزایش یافته و به حدود در حدود 40kb می رسد.
امکان محاسبه تعداد کلون هایی که باید در یک کتابخانه ژنومی حضور داشته باشد و احتمال حضور یک توالی DNA خاص به وسیله فرمول زیر به دست می آید:
N= In(1-P)/ In(1-f)
N تعداد نوترکیب ها بوده و P احتمال مورد نظر و f نسبت طول قطعات كلون شونده به طول ژنوم می باشد.
به عنوان مثال با احتمال 99% و با اندازه قطعه وارد شونده 20kb، تعداد نوترکیب ها برای اشریشیا کولی با طولی برابر ۱۰۶×۴٫۶ برابر است با:= 1.1x103
/In(1- 0.99)
In [1-(2x104/4.6×100)]
مفهوم کتابخانه ژنی (Gene Library) – که گاهی با نامهای کتابخانه DNA یا کتابخانه اسید نوکلئیک (Nucleic-Acid Library) نیز شناخته میشود – پایه و اساس زیستمولکولی و ژنومیک مدرن است. یک کتابخانه ژنی، مجموعهای سیستماتیک و نماینده از قطعات DNA یا cDNA (Complementary DNA – دیانای مکمل) است که بهطور کامل اطلاعات ژنتیکی یک ارگانیسم، بافت یا نمونه محیطی را دربرمیگیرد.
این قطعات در قالبی پایدار نگهداری میشوند—بهطور کلاسیک در وکتورهای کلونینگ (Cloning Vectors) مانند پلاسمیدها (Plasmids) یا باکتریوفاژها (Bacteriophages) و در روشهای جدیدتر به صورت مولکولهای دارای آداپتور (Adaptor-Ligated Molecules) سازگار با پلتفرمهای توالییابی نسل جدید (Next-Generation Sequencing – NGS)—تا پژوهشگران بتوانند آنها را بارها بازیابی، تکثیر و تحلیل کنند. ایجاد چنین مجموعهای، منبعی تجدیدپذیر در اختیار دانشمندان قرار میدهد که میتواند کل ژنوم یا بخش بیانشده آن را در یک زمان مشخص بهطور کامل نمایش دهد.
دیدگاه تاریخی (Historical Perspective)
ریشههای کتابخانههای ژنی به دههٔ ۱۹۷۰ برمیگردد؛ زمانی که فناوری DNA نوترکیب (Recombinant DNA Technology) برای نخستین بار به دانشمندان امکان داد قطعات تعریفشدهٔ ژنوم را جدا و تکثیر کنند.
-
روشهای اولیه: پیشگامان این حوزه از آنزیمهای برشدهنده (Restriction Enzymes) برای خرد کردن ژنومهای بزرگ به قطعات قابل مدیریت استفاده کردند و سپس این قطعات را در وکتورهای باکتریایی (Bacterial Vectors) وارد نمودند و بدین ترتیب نخستین کتابخانههای ژنومی (Genomic Libraries) شکل گرفت.
-
این کتابخانهها برای نقشهبرداری ژنهای مرتبط با بیماریهای انسانی، شناسایی عناصر تنظیمی و ساخت توالیهای کامل ژنومهای باکتریایی و یوکاریوتی ضروری بودند.
-
دستاوردهای بزرگی مانند پروژه ژنوم انسان (Human Genome Project) به شدت به کتابخانههای با درج بزرگ (Large-Insert Libraries) از جمله کروموزومهای مصنوعی باکتریایی (BAC: Bacterial Artificial Chromosome) و کروموزومهای مصنوعی مخمری (YAC: Yeast Artificial Chromosome) متکی بودند که قطعات همپوشان برای نقشهبرداری فیزیکی و توالییابی فراهم میکردند.
با پیشرفت شیمی و ابزارهای توالییابی (Sequencing Chemistry & Instrumentation)، روشهای تهیه کتابخانه از کلونینگ پرزحمت به فرآیندهای آنزیمی کارآمد و پرظرفیت برای پلتفرمهای NGS تغییر یافت. امروزه به جای وارد کردن قطعات به سلولهای زنده، رویکردهای مدرن آداپتورهای اختصاصی پلتفرم را به DNA برشخورده یا cDNA مشتقشده از RNA متصل میکنند و امکان توالییابی میلیاردها مولکول به صورت موازی را فراهم میسازند.
با وجود این جهشهای فناورانه، اصل بنیادین همچنان یکسان است: کتابخانه ژنی مجموعهای مشخص و بازتولیدپذیر از قطعات اسید نوکلئیک است که بازتابدهنده دقیق یک ژنوم یا ترنسکریپتوم (Transcriptome – مجموعه کامل RNAهای بیانشده) میباشد.
منطق اصلی و اهمیت علمی (Core Rationale and Scientific Importance)
ایجاد کتابخانه ژنی چندین هدف کلیدی را دنبال میکند:
-
نمایش جامع ماده ژنتیکی: تضمین میکند که توالیهای نادر، نواحی تنظیمی و واریانتهای ساختاری (Structural Variants) همگی برای آنالیز حفظ شوند.
-
منبع استاندارد و تجدیدپذیر: یک مجموعه پایدار که میتواند میان آزمایشگاهها به اشتراک گذاشته شود یا حتی سالها بعد با فرضیهها و فناوریهای جدید دوباره تحلیل شود.
-
کاوش هدفمند پرسشهای خاص:
-
کتابخانه cDNA میتواند نشان دهد کدام ژنها در یک بافت سرطانی بهطور فعال بیان میشوند.
-
کتابخانه متاژنومیک (Metagenomic Library) تنوع میکروبی یک نمونه خاک را بدون نیاز به کشت میکروارگانیسمها آشکار میکند.
-
-
کاربردهای پزشکی:
-
کتابخانههای ژنومی کامل امکان شناسایی واریانتهای ساختاری مرتبط با بیماریهای ژنتیکی را میدهند.
-
کتابخانههای RNA-seq امضای بیان ژنی (Expression Signatures) را آشکار میسازند که به انتخاب درمان سرطان کمک میکند.
-
کتابخانههای محیطی و متاژنومیک ردگیری ژنهای مقاومت آنتیبیوتیکی را در اکوسیستمها ممکن میکنند و بهعنوان هشدار زودهنگام برای تهدیدهای بهداشت عمومی عمل مینمایند.
-
در پژوهش بنیادی، این کتابخانهها برای ژنومیک عملکردی (Functional Genomics)، ژنومیک مقایسهای (Comparative Genomics)، مطالعات تکاملی (Evolutionary Studies) و کشف آنزیمها یا مسیرهای متابولیکی جدید حیاتی هستند.
چارچوب مفهومی (Conceptual Framework)
با وجود آنکه اصطلاح کتابخانه ژنی ممکن است یک مخزن ساده را تداعی کند، ساخت آن یک فرآیند چندمرحلهای دقیق است که نیاز به حفظ کیفیت اسید نوکلئیک و طراحی آزمایشی محتاطانه دارد.
-
چه هدف، ساخت یک کتابخانه ژنومی جامع (Comprehensive Genomic Library) باشد، چه یک کتابخانه cDNA متمرکز بر ترنسکریپت (Transcript-Focused cDNA Library) یا پنل توالییابی هدفمند (Targeted Sequencing Panel)، گام نخست همیشه استخراج DNA یا RNA با کیفیت بالا از ماده منبع است.
-
مراحل بعدی شامل برش کنترلشده (Controlled Fragmentation)، ترمیم انتها (End Repair)، اتصال آداپتور (Adaptor Ligation) و در صورت نیاز تکثیر (Amplification) است تا یک مخزن پایدار از قطعات سازگار با آنالیز یا توالییابی ایجاد شود.
هر یک از این مراحل بر نمایندگی و کیفیت نهایی کتابخانه تأثیر دارد و بنابراین، آمادهسازی کتابخانه یک عامل تعیینکننده موفقیت آزمایش است.
تحول به سمت کاربردهای پرظرفیت (Evolution Toward High-Throughput Applications)
ظهور توالییابی نسل جدید (Next-Generation Sequencing – NGS) فرآیند آمادهسازی کتابخانه ژنی را از یک تکنیک محدود کلونینگ به دروازه اصلی ژنومیک تبدیل کرده است.
-
پلتفرمهای NGS مانند Illumina، Pacific Biosciences (PacBio) و Oxford Nanopore هر کدام به استراتژی ساخت کتابخانه مختص خود نیاز دارند تا با طول خوانش (Read Length)، پروفایل خطا (Error Profile) و توان عملیاتی (Throughput) سازگار باشند.
-
اتوماسیون (Automation)، میکروفلوئیدیک (Microfluidics) و روش تگمنتیشن (Transposase-Mediated Tagmentation) اکنون امکان آمادهسازی صدها یا هزاران کتابخانه به صورت موازی را فراهم کردهاند، که منجر به کاهش چشمگیر هزینه و زمان در عین حفظ دقت بالا شده است.
-
توالییابی تکسلولی (Single-Cell Sequencing) این حوزه را حتی فراتر برده و نیازمند پروتکلهای با ورودی بسیار کم (Ultra-Low-Input Protocols) و بارکدگذاری مولکولی دقیق (Molecular Barcoding) برای ثبت محتوای ژنومی یا ترنسکریپتومی هر سلول منفرد است.
۲.۱ کتابخانههای ژنومی (Genomic Libraries)
یک کتابخانه ژنومی شامل قطعاتی است که بهصورت جمعی نمایانگر کل ژنوم یک ارگانیسم میباشند؛ این شامل ناحیههای کدکننده و غیرکدکننده، عناصر تنظیمی، اینترونها و توالیهای تکراری است. هدف این روش، حفظ تمام اطلاعات ژنومی در یک قالب پایدار و قابل کلون شدن است.
-
ساختار و روش تهیه: در گذشته، ساخت این کتابخانهها با استفاده از DNA با وزن مولکولی بالا آغاز میشد که بهطور جزئی توسط آنزیمهای برشدهنده (restriction enzymes) هضم یا بهصورت مکانیکی به قطعاتی با اندازه مشخص شکسته میگردید. سپس این قطعات به وکتورهایی مانند لامبدا فاژ (λ phage)، کاسمید (cosmid)، کروموزومهای مصنوعی باکتریایی (BACs) یا کروموزومهای مصنوعی مخمری (YACs) متصل میشدند. برای مثال، کتابخانههای BAC قادر به نگهداری قطعاتی با اندازه ۱۰۰ تا ۳۰۰ کیلوباز هستند که آنها را برای پروژههای عظیمی مثل پروژه ژنوم انسان (Human Genome Project) ارزشمند میکند.
-
مزایا:
-
کامل بودن و پایداری بالا
-
حفظ عناصر غیرکدکننده مانند پروموترها و اینترونها
-
مناسب برای مطالعات تنظیم بیان ژن، ژنومیک ساختاری و ژنومیک تطبیقی
-
-
نکته مهم: امروزه پلتفرمهای توالییابی نسل جدید (NGS) میتوانند دادههایی حتی غنیتر از روشهای کلاسیک کلونینگ تولید کنند، با این حال مفهوم نمایش کامل ژنوم در قالبی قابل مدیریت همچنان اساسی است، بهویژه در توالییابی طولانی (long-read) و متاژنومیک.
۲.۲ کتابخانههای cDNA
کتابخانههای cDNA (Complementary DNA) اساساً با کتابخانههای ژنومی تفاوت بنیادی دارند، زیرا تنها شامل ژنهای بیانشده در یک سلول یا بافت در زمان مشخص هستند.
-
روش ساخت: ابتدا RNA پیامرسان (mRNA) استخراج میشود (معمولاً با پرایمرهای Oligo(dT) که به دُم پلی(A) متصل میشوند). سپس آنزیم ترانسکریپتاز معکوس (Reverse Transcriptase) یک رشته cDNA سنتز میکند که بعداً به DNA دو رشتهای تبدیل و کلون یا مستقیماً برای توالییابی آماده میشود.
-
ویژگیها:
-
فاقد اینترونها است زیرا فرآیند Splicing در mRNA قبلاً انجام شده است.
-
بازتابی مستقیم از ترنسکریپتوم (Transcriptome) سلول است.
-
-
کاربردها:
-
کشف ژنها، بررسی Alternative splicing، پروفایل بیان ژن
-
مطالعات زیستشناسی تکوینی (مثلاً بررسی مراحل مختلف جنینی)
-
پژوهشهای بالینی برای شناسایی آنکوژنها یا بیومارکرهای تومور
-
-
نکته مهم: بیان ژن وابسته به شرایط است؛ بنابراین دو کتابخانه cDNA از یک ارگانیسم میتوانند بر اساس نوع بافت یا شرایط محیطی بسیار متفاوت باشند. ارزیابی دقیق کیفیت RNA برای جلوگیری از بایاس بسیار حیاتی است.
۲.۳ کتابخانههای متاژنومیک (Metagenomic Libraries)
کتابخانههای متاژنومیک مستقیماً از DNA استخراجشده از جوامع میکروبی پیچیده و بدون نیاز به کشت گونههای منفرد ساخته میشوند.
-
اهمیت: این روش باعث انقلاب در شناخت ما از میکروبیومها در خاک، آب اقیانوس و روده انسان شده است.
-
کاربردها:
-
کشف آنزیمهای جدید، مسیرهای متابولیکی و ژنومهای میکروارگانیسمهای ناشناخته
-
غربالگری عملکردی برای شناسایی کلونهای دارای آنزیمهای صنعتی (مانند سلولاز یا لیپاز)
-
تحلیل تنوع جمعیت میکروبی و تعاملات اکولوژیک
-
-
نکته کلیدی: بسیاری از میکروبها قابل کشت نیستند، بنابراین متاژنومیک تنها راه مشاهده تنوع پنهان زمین و ظرفیت آن برای زیستفناوری است.
۲.۴ کتابخانههای BAC و YAC
برای پروژههایی که نیاز به قطعات DNA بسیار بزرگ دارند از کتابخانههای BAC (Bacterial Artificial Chromosome) و YAC (Yeast Artificial Chromosome) استفاده میشود.
-
BAC:
-
مشتق از پلاسمید F در E. coli
-
ظرفیت ۱۰۰–۳۰۰ کیلوباز
-
پایداری بالا
-
مناسب برای نقشهبرداری فیزیکی، کلونینگ موضعی و مونتاژ ژنومهای پیچیده
-
-
YAC:
-
قابلیت حمل تا ۱ مگاباز DNA
-
با این حال مستعد بازآرایی (rearrangement) بوده و امروزه بیشتر با BAC جایگزین شده است.
-
-
کاربرد مشترک: بسیار مفید برای توالییابی ژنومهای یوکاریوتی پیچیده، مطالعات ساختاری و بستن شکافهای اسمبلی.
۲.۵ کتابخانههای فوزمید (Fosmid Libraries)
فوزمیدها وکتورهای هیبریدی هستند که ویژگیهای پلاسمید و فاژ را ترکیب میکنند و معمولاً ۳۰–۴۰ کیلوباز DNA را حمل میکنند.
-
ویژگیها:
-
پایداری بالاتر از کاسمیدها
-
مناسب برای توالییابی ناحیههای تکراری یا GC بالا که در BAC ممکن است ناپایدار باشند.
-
-
کاربرد: برای اعتبارسنجی اسمبلی ژنوم یا ساخت پنل مرجع برای آنالیز تغییرات ساختاری استفاده میشود.
۲.۶ کتابخانههای بیانی (Expression Libraries)
کتابخانه بیانی نوعی کتابخانه cDNA یا ژنومی است که توالیهای درجشده در آن زیر پروموترهای قوی کلون میشوند تا در میزبان رونویسی و ترجمه شوند.
-
هدف: تولید پروتئینها یا پپتیدهای فعال برای غربالگری فعالیتها یا برهمکنشها.
-
کاربرد:
-
کشف آنتیبادی
-
نقشهبرداری اپیتوپ
-
شناسایی برهمکنشهای پروتئین–پروتئین یا پروتئین–DNA
-
-
نمونه بارز: کتابخانههای فاژ دیسپلی (Phage Display) که در آن پپتیدها یا آنتیبادیها بر سطح باکتریوفاژ نمایش داده میشوند.
۲.۷ کتابخانههای هدفمند یا سفارشی (Targeted or Custom Libraries)
همه کتابخانهها به دنبال پوشش کامل ژنوم نیستند.
-
هدف: غنیسازی برای نواحی ژنومی یا مجموعه ژنهای خاص با استفاده از پروبهای هیبرید کپچر یا PCR چندگانه.
-
کاربرد بالینی:
-
پنلهای ژنی برای بیماریهای سرطانی ارثی
-
کاردیومیوپاتی
-
ویژگیهای فارماکوژنتیکی
-
این روش باعث افزایش کارایی توالییابی و تمرکز بر نواحی کلیدی با عمق و دقت بالا میشود.
۲.۸ کتابخانههای تکسلولی (Single-Cell Libraries)
ژنومیک و ترنسکریپتومیک تکسلولی نیازمند روشهای فوق حساس است که قادر به دریافت اسید نوکلئیک از یک سلول منفرد باشند.
-
روشها: استفاده از دستگاههای میکروفلوئیدیک یا روشهای قطرهای (Droplet-based) برای جداسازی سلولها و افزودن بارکدهای مولکولی که هر خوانش را به سلول منشاء متصل میکند.
-
کاربرد:
-
بررسی ناهمگنی سلولی در بافتهایی مانند تومور یا جنین در حال رشد
-
شناسایی زیرجمعیتها و سلولهای نادر که در آنالیز جمعی پنهان میمانند.
-