چگونه سیگنالهای کلسیمی غیرعصبی واحدهای نوری چشم را هماهنگ میکنند؟
نقش موجهای کلسیمی غیرعصبی در شکلگیری و همراستایی واحدهای نوری چشم مگس سرکه
خلاصه
چشم مرکب مگس سرکه نمونهای برجسته از خودسازماندهی سلولی در طی تکوین بافتی است. در این پژوهش نشان داده میشود که سلولهای پشتیبان غیرعصبی شبکیه، از طریق تولید موجهای کلسیمی خودبهخودی، نقشی اساسی در تنظیم معماری دقیق چشم ایفا میکنند. این موجها با فعالسازی گیرنده تیروزینکینازی Cad96Ca آغاز شده و از مسیر پیامرسانی PLC-γ–IP3R موجب آزادسازی کلسیم از شبکه آندوپلاسمی میشوند. سپس، یک «کد اینکسینی» خاص، انتشار فضایی موجها را از طریق اتصالات شکافدار میان سلولی هدایت میکند. شدت این موجها با اندازه واحدهای نوری (اوماتیدیا) همبستگی داشته و موجب انقباضات آپیکال متناسب در مرزهای بین واحدی میشود. این فرایند، همراستایی دقیق لنزها و سازمانیافتگی منظم شبکیه را تضمین میکند. یافتهها نشان میدهند که فعالیت هماهنگ کلسیمی در سلولهای غیرعصبی، یکی از سازوکارهای بنیادین در الگودهی بافتی سامانه بینایی در مرحله رشد است.
مقدمه
کارکرد صحیح سامانههای حسی مستلزم سازمانیافتگی دقیق اجزای سلولی آنهاست. چشم، بهعنوان یکی از پیچیدهترین اندامهای حسی، نمونهای شاخص از این اصل بهشمار میرود؛ جایی که جایگیری دقیق نورونها و سلولهای پشتیبان برای دریافت و پردازش مؤثر نور ضروری است. با این حال، سازوکارهای سلولی که این هماهنگی فضایی را در طی تکوین ایجاد میکنند، هنوز بهطور کامل شناخته نشدهاند.
در بسیاری از سیستمهای زیستی، موجهای کلسیمی هماهنگ در نورونها نقش کلیدی در شکلگیری مدارهای عصبی ایفا میکنند. برای مثال، در شبکیه پستانداران، موجهای کلسیمی اولیه مسیرهای ارتباطی میان چشم و مغز را هدایت میکنند. با این وجود، نقش موجهای کلسیمی در سلولهای غیرعصبی و سهم آنها در سازماندهی معماری بافتی چشم تاکنون کمتر مورد توجه قرار گرفته است.
شبکیه مگس سرکه (Drosophila melanogaster) به دلیل ساختار منظم و تکرارشونده خود، مدل مناسبی برای بررسی این پدیده محسوب میشود. چشم مرکب این حشره از حدود ۸۰۰ واحد نوری (اوماتیدیا) تشکیل شده است که هر یک شامل هشت نورون گیرنده نور بوده و توسط مجموعهای از سلولهای پشتیبان غیرعصبی با ویژگیهای گلیایی و اپیتلیالی احاطه میشوند. این آرایش منظم، بستری ایدهآل برای مطالعه تعاملات سلولی و الگوهای هماهنگسازی در مقیاس بافتی فراهم میکند.
نتایج
بروز موجهای کلسیمی اولیه در شبکیه در حال تکوین
با بهرهگیری از تصویربرداری زنده و نشانگر فلورسنت GCaMP6s، دینامیک کلسیم در سلولهای شبکیه در طول رشد بررسی شد. دو بازه زمانی متمایز از فعالیت موجی کلسیم شناسایی گردید:
-
موجهای اولیه که حدود ۳۷ ساعت پس از آغاز تکوین چشم ظاهر شده و تقریباً ۳ ساعت دوام دارند.
-
موجهای ثانویه که از حدود ۷۳ ساعت پس از تکوین آغاز شده و تا زمان خروج حشره بالغ ادامه مییابند.
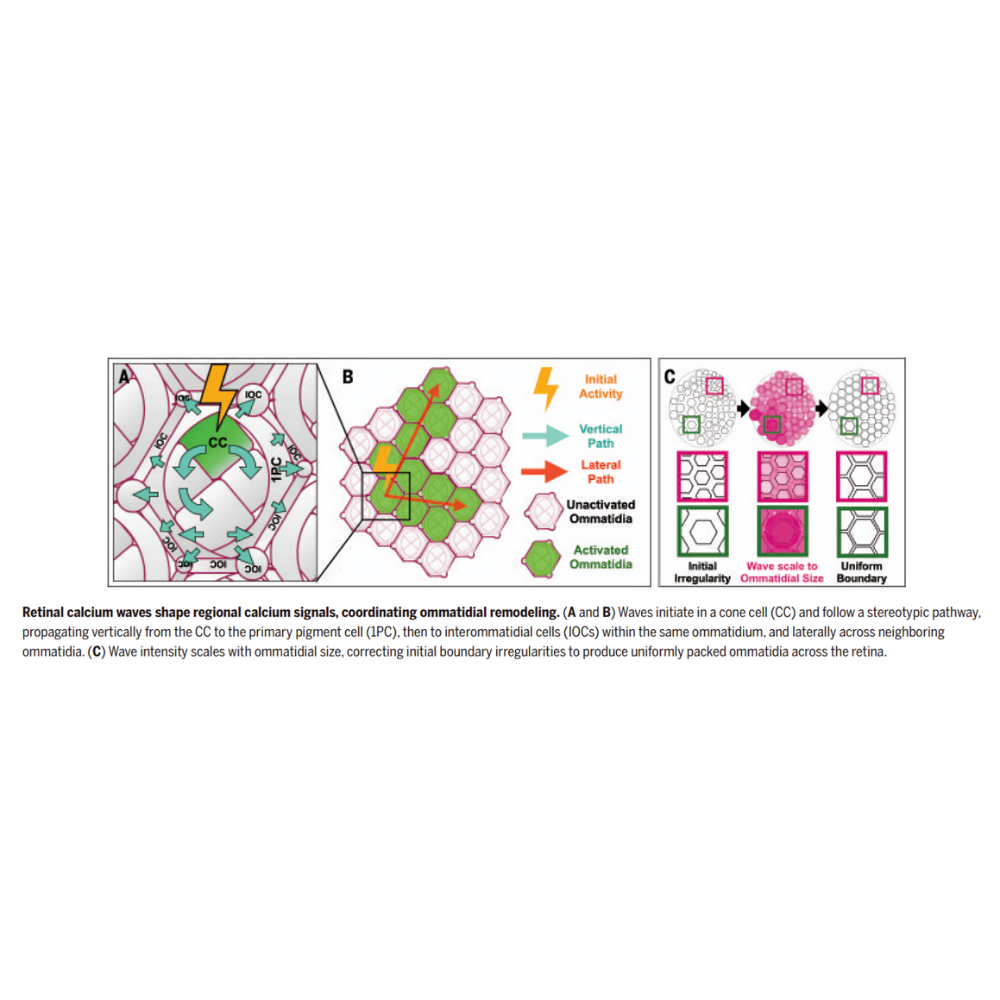
موجهای اولیه معمولاً از یک سلول مخروطی آغاز شده و سپس به سلولهای غیرعصبی مجاور در همان واحد نوری گسترش مییابند. نورونهای گیرنده نور در این انتشار مشارکت مستقیمی ندارند، بلکه این سلولهای پشتیبان هستند که شبکهای هماهنگ و منظم را شکل میدهند.
مسیر پیامرسانی Cad96Ca–PLCγ–IP3R
آغاز موجهای کلسیمی به فعالسازی گیرنده Cad96Ca در سلولهای مخروطی وابسته است. این فعالسازی، مسیر PLC-γ را تحریک کرده و منجر به تولید اینوزیتولتریفسفات (IP3) میشود . IP3 با اثر بر گیرندههای خود در شبکه آندوپلاسمی، آزادسازی کلسیم درونسلولی را القا میکند. دستکاریهای ژنتیکی نشان دادند که اختلال در هر یک از اجزای این مسیر، آغاز و انتشار موجهای کلسیمی را بهطور جدی مختل میکند.
نقش اینکسینها و اتصالات شکافدار
انتشار فضایی موجها از طریق شبکهای از اتصالات شکافدار (Gap Junctions) انجام میشود که توسط پروتئینهای اینکسین تشکیل شدهاند. اگرچه انواع مختلفی از اینکسینها در سلولهای غیرعصبی شبکیه بیان میشوند، ترکیب خاصی از Inx1، Inx2 و Inx3 مسئول هدایت مؤثر موجهاست. Inx2 برای انتشار کلی موجها ضروری است، در حالی که Inx3 نقشی ویژه در انتقال سیگنال میان سلولهای بین واحدی ایفا میکند.
انقباضات آپیکال وابسته به اندازه واحدهای نوری
تحلیل شدت موجها نشان داد که در نواحی دارای اوماتیدیای بزرگتر، دامنه موجهای کلسیمی افزایش مییابد. این افزایش شدت، موجب انقباضات آپیکال قویتر در مرزهای بین واحدهای نوری شده و در نتیجه، همراستایی و یکپارچگی بافت شبکیه حفظ میشود. این سازوکار نقش مهمی در شکلگیری دقیق لنزها ایفا میکند.
پیامدهای ساختاری بلندمدت
موجهای کلسیمی نهتنها بر سازماندهی کلی شبکیه اثر میگذارند، بلکه برای شکلگیری صحیح ساختارهای داخلی لنز، از جمله شبهمخروطها (pseudocones) نیز ضروری هستند. در جهشهایی که این موجها مختل میشوند، این ساختارهای داخلی ناقص باقی میمانند، حتی در شرایطی که ظاهر بیرونی لنز طبیعی به نظر میرسد.
بحث و نتیجهگیری
یافتههای این پژوهش نشان میدهد که سلولهای پشتیبان غیرعصبی شبکیه، از طریق تولید و هماهنگسازی موجهای کلسیمی، نقشی فعال و تعیینکننده در تکوین معماری دقیق چشم ایفا میکنند. این موجها پیش از آغاز فعالیت نورونی رخ داده، توسط Cad96Ca آغاز میشوند و انتشار آنها از طریق شبکهای سازمانیافته از اینکسینها هدایت میگردد. همبستگی شدت موجها با اندازه واحدهای نوری، امکان ایجاد انقباضات متناسب و تنظیم دقیق مرزهای بافتی را فراهم میسازد.
به بیان ساده، این موجهای کلسیمی همچون یک سامانه هدایتکننده عمل میکنند که سلولهای پشتیبان را بهصورت هماهنگ در زمان و فضا تنظیم کرده و زمینهساز شکلگیری ساختاری منظم و کارآمد برای عملکرد بینایی میشوند. این نتایج، درک ما را از نقش سلولهای غیرعصبی در الگودهی بافتی گسترش داده و افقهای تازهای برای مطالعه تکوین اندامهای حسی میگشاید.
منابع و توضیحات:
Ben Jiwon Choi et al.,Retinal calcium waves coordinate uniform tissue patterning of the Drosophila eye.Science390,eady5541(2025). DOI:10.1126/science.ady5541