
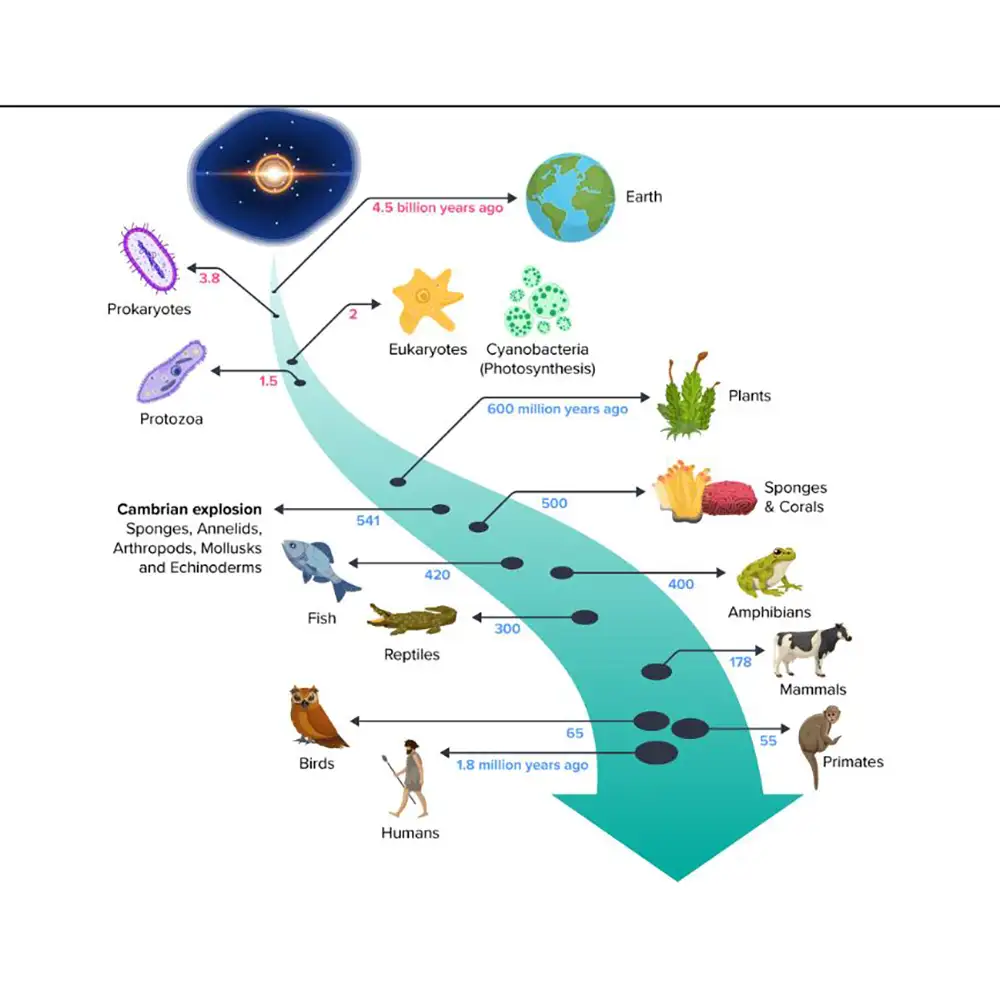
تکامل چیست؟
تعریف تکامل (Defining Evolution)
در علم زیستشناسی (Biological Science)، تکامل (Evolution) به معنای تغییرات وراثتی (Heritable Change) در جمعیتهای موجودات زنده در طول نسلهای پیدرپی است. تکامل تنها توصیفی از تغییر نیست، بلکه یک چارچوب تبیینی قدرتمند (Robust Explanatory Framework) است که هم تنوع شگفتانگیز حیات (Astonishing Diversity of Life) و هم وحدت عمیق بین همهٔ ارگانیسمهای زمین (Deep Unity of Organisms) را توضیح میدهد. نظریهٔ تکامل (Evolutionary Theory) تشریح میکند که چگونه مولکولهای ساده میلیاردها سال پیش به واحدهای خودتکثیرشونده (Self-Replicating Entities) تبدیل شدند و چگونه این واحدها، از طریق ترکیبی از تنوع ژنتیکی تصادفی (Random Genetic Variation) و فرآیندهای نظاممند مانند انتخاب طبیعی (Natural Selection)، سرانجام منجر به ایجاد تنوع عظیم گیاهان، جانوران، قارچها و میکروبهایی شدند که امروزه مشاهده میکنیم. در اصل، تکامل مطالعهٔ «تبار با تغییر» (Descent with Modification) است که تغییرات در سطح ژنها، صفات، جمعیتها و حتی کل اکوسیستمها را در بر میگیرد.
ریشههای تاریخی (Historical Foundations)
ریشههای فکری مفهوم تکامل به دوران باستان بازمیگردد. فیلسوفان یونانی اولیه مانند آناکسیماندر (Anaximander) گمان میکردند که حیات از آب پدید آمده و به تدریج دگرگون شده است، در حالی که اندیشمندانی مانند لوکرتیوس (Lucretius) جهانی را توصیف کردند که توسط علل طبیعی شکل گرفته است نه مداخلهٔ ماوراءطبیعی. در قرون بعد، طبیعتشناسان با مشاهدهٔ فسیلها (Fossils) و شباهتهای آناتومیکی بین گونهها، به تاریخ پویا و متغیر حیات پی بردند؛ اما این مشاهدات فاقد یک سازوکار یکپارچه بود. این وضعیت در قرن نوزدهم با کارهای انقلابی چارلز داروین (Charles Darwin) و آلفرد راسل والاس (Alfred Russel Wallace) تغییر کرد. این دو دانشمند به طور مستقل انتخاب طبیعی (Natural Selection) را به عنوان عامل اصلی تغییر تکاملی شناسایی کردند. بینشهای آنها که در سال ۱۸۵۸ ارائه شد و در کتاب مشهور داروین «خاستگاه گونهها» (On the Origin of Species) در سال ۱۸۵۹ گسترش یافت، توضیحی آزمونپذیر برای انطباق (Adaptation) و تنوع زیستی (Diversification) ارائه داد.
اوایل قرن بیستم شاهد شکلگیری سنتز مدرن (Modern Synthesis) بود؛ ادغامی مهم از انتخاب طبیعی داروینی (Darwinian Natural Selection) با ژنتیک مندل (Mendelian Genetics). دانشمندانی مانند رونالد فیشر (Ronald Fisher)، ج. ب. اس. هالدین (J. B. S. Haldane) و سیوال رایت (Sewall Wright) نشان دادند که جهش (Mutation)، بازترکیبی (Recombination) و رانش ژنتیکی (Genetic Drift) چگونه درون جمعیتها عمل میکنند و تغییر تکاملی ایجاد مینمایند. این سنتز نه تنها ایدههای مرکزی داروین را تأیید کرد، بلکه آنها را گسترش داد و ژنتیک جمعیت (Population Genetics) را به عنوان ستون ریاضی زیستشناسی تکاملی تثبیت نمود.
نقش محوری در زیستشناسی مدرن (Central Role in Modern Biology)
نظریهٔ تکامل اصل سازماندهنده (Organizing Principle) علوم زیستی است. رشتههایی چون ژنتیک (Genetics)، زیستشناسی تکوینی (Developmental Biology)، بومشناسی (Ecology) و دیرینشناسی (Paleontology) همگی در چارچوب تکاملی برای توضیح الگوهای طبیعی همگرا میشوند. درک تکامل به پژوهشگران کمک میکند سابقهٔ فسیلی (Fossil Record) را تفسیر کنند، تبار گونهها را ردیابی نمایند و پیشبینی کنند که ارگانیسمها چگونه به فشارهای محیطی پاسخ خواهند داد. تکامل اساس فیلوژنتیک (Phylogenetics) است که با استفاده از ویژگیهای ریختی و دادههای مولکولی، تاریخ شاخهبندی حیات را بازسازی میکند. همچنین در حفاظت زیستمحیطی (Conservation Biology) با تأکید بر اهمیت تنوع ژنتیکی (Genetic Diversity) برای بقا نقش اساسی دارد.
فراتر از پژوهش پایه، تکامل در حوزههای کاربردی نیز حیاتی است. در پزشکی، تکامل سریع پاتوژنها (Rapid Evolution of Pathogens) ظهور مقاومت آنتیبیوتیکی (Antibiotic Resistance) را توضیح میدهد و به توسعهٔ درمانها و واکسنها جهت میدهد. زیستشناسی سرطان (Cancer Biology) اکنون تومورها را به عنوان جمعیتهای در حال تکامل سلولی (Evolving Populations of Cells) میبیند؛ دیدگاهی که راهکارهای درمانی برای به تأخیر انداختن یا جلوگیری از مقاومت دارویی را شکل میدهد. در کشاورزی، انتخاب مصنوعی (Artificial Selection) که همان پرورش آگاهانهٔ گونهها توسط انسان است، کاربرد مستقیم اصول تکاملی است و منجر به تولید محصولات و دامهای امروزی شده است. حتی در علوم کامپیوتر (Computer Science)، الگوریتمهای تکاملی (Evolutionary Algorithms) با تقلید از انتخاب طبیعی مسائل پیچیدهٔ بهینهسازی را حل میکنند.
فرآیندها و سازوکارهای کلیدی (Key Processes and Mechanisms)
تکامل از طریق چند سازوکار شناختهشده (Well-Characterized Mechanisms) عمل میکند:
-
جهش (Mutation): ایجاد تنوع ژنتیکی جدید با تغییر توالی DNA.
-
انتخاب طبیعی (Natural Selection): گزینش صفاتی که بقا و تولیدمثل را بهبود میبخشند.
-
رانش ژنتیکی (Genetic Drift): نوسان تصادفی فراوانی اللها بهویژه در جمعیتهای کوچک.
-
جریان ژن (Gene Flow/Migration): جابجایی اللها میان جمعیتها که میتواند صفات سودمند را گسترش دهد یا سازگاری محلی را خنثی کند.
این فرآیندها با هم الگوهای انطباق (Adaptation)، واگرایی (Divergence) و گاه انقراض (Extinction) را در سراسر درخت حیات (Tree of Life) ایجاد میکنند. تکامل هیچ هدف از پیش تعیینشدهای ندارد (No Predetermined Goal)؛ بلکه نتیجهٔ برهمکنش پویای شانس (Chance) و ضرورت (Necessity) است که در آن جهشهای تصادفی و رویدادهای تاریخی با فشارهای انتخابی مداوم تعامل میکنند.
شواهد تکامل (Evidence Base)
شواهد پشتیبان تکامل وسیع و چندجانبه است:
-
سابقهٔ فسیلی (Fossil Record) تغییرات تدریجی در زمان زمینشناسی و شکلهای گذار مانند دایناسورهای پردار (Feathered Dinosaurs) را که خزندگان را به پرندگان مدرن پیوند میدهند، نشان میدهد.
-
آناتومی مقایسهای (Comparative Anatomy) ساختارهای همولوگ (Homologous Structures) مانند استخوانهای اندام در نهنگ، خفاش و انسان را آشکار میکند که نشاندهندهٔ نیای مشترک (Common Ancestry) است.
-
جنینشناسی (Embryology) مسیرهای تکوینی حفظشده در گونههای مختلف را نمایان میسازد.
-
در سطح مولکولی، جهانی بودن کد ژنتیکی (Universality of the Genetic Code) و وجود توالیهای مشترک در DNA و پروتئینها روابط تکاملی عمیق را نشان میدهد.
-
زیستجغرافیا (Biogeography) نیز تأیید بیشتری میافزاید؛ گونههای جزیرهای غالباً به فرمهای سرزمین اصلی شباهت دارند اما سازگاریهای منحصربهفرد (Unique Adaptations) ناشی از انزوا را نشان میدهند.
اهمیت گسترده (Broader Significance)
تأثیر تفکر تکاملی فراتر از زیستشناسی است و بر فلسفه (Philosophy)، آموزش (Education) و سیاستگذاری عمومی (Public Policy) اثر میگذارد. این نظریه مباحثی چون حفاظت تنوع زیستی (Biodiversity Conservation)، سازگاری با تغییرات اقلیمی (Climate Change Adaptation) و اخلاق مهندسی ژنتیک (Ethics of Genetic Engineering) را شکل میدهد. همچنین درک جایگاه بشر به عنوان بخشی از یک تبار پیوسته (Continuous Lineage) و نه موجودی جدا از بقیهٔ حیات، احترام عمیقتری به ارتباطات اکوسیستمی (Interconnectedness of Ecosystems) و مسئولیت حفظ آنها القا میکند.
۲. بنیانهای نظریهٔ تکامل (Foundations of Evolutionary Theory)
۲.۱ تنوع و وراثت: مواد خام تکامل (Variation and Heredity: The Raw Materials of Evolution)
در قلب تکامل (Evolution)، تنوع ژنتیکی (Genetic Variation) قرار دارد؛ یعنی تفاوتهای توالی DNA میان افراد یک جمعیت. این تنوع **مواد خام (Raw Material)**ی است که نیروهای تکاملی بر آن اثر میگذارند. این تفاوتها از چندین منبع ایجاد میشوند:
-
جهشها (Mutations) که تغییرات در توالی DNA هستند و میتوانند بر اثر خطاهای همانندسازی، تماس با تابش (Radiation) یا مواد شیمیایی، یا فعالیت عناصر ژنتیکی متحرک به وجود آیند. بسیاری از جهشها خنثی یا زیانآورند، اما برخی مزایایی ایجاد میکنند که بقا (Survival) یا موفقیت تولیدمثلی (Reproductive Success) را افزایش میدهد.
-
بازترکیبی (Recombination)، یعنی درهمآمیختن مادهٔ ژنتیکی در طول میوز (Meiosis) که ترکیبهای جدیدی از اللها (Alleles) میسازد و تنوع فنوتیپی را افزایش میدهد.
حتی در موجودات تکیاختهای با تولیدمثل غیرجنسی (Asexual Reproduction)، مکانیسمهایی مانند انتقال افقی ژن (Horizontal Gene Transfer) عناصر ژنتیکی جدیدی وارد میکنند.
برای آنکه تغییر تکاملی پایدار بماند، این تنوع باید قابل وراثت (Heritable) باشد. آزمایشهای گرگور مندل (Gregor Mendel) روی گیاهان نخود در سدهٔ نوزدهم نشان داد که صفات از طریق واحدهای مجزا (Discrete Units) – که امروزه ژن (Gene) نامیده میشوند – منتقل میشوند و نه با هم مخلوط. بازکشف کارهای مندل در اوایل قرن بیستم، درک مکانیکی وراثت (Heredity) را با انتخاب طبیعی داروین (Darwinian Natural Selection) ادغام کرد. امروز، زیستشناسی مولکولی (Molecular Biology) و ژنومیک (Genomics) تأیید میکنند که DNA دستورالعملهای ساخت موجودات زنده را رمزگذاری میکند و این دستورها در نسلهای متوالی تقریباً بهدقت کپی میشوند؛ هرچند خطاهای گاهبهگاه، منبعی برای نوآوری تکاملیاند.
۲.۲ انتخاب طبیعی: موتور سازگاری (Natural Selection: The Engine of Adaptation)
انتخاب طبیعی (Natural Selection) سازوکار اصلی است که داروین برای توضیح سازگار شدن موجودات با محیطشان شناسایی کرد. این فرایند هنگامی رخ میدهد که سه شرط برقرار باشد:
-
افراد از نظر صفات متفاوت (Variation in Traits) باشند،
-
آن صفات بر بقا یا تولیدمثل (Survival or Reproduction) اثر بگذارند،
-
و آن صفات قابل وراثت (Heritable) باشند.
در چنین حالتی، افرادی که ویژگیهای سودمند دارند، فرزندان بیشتری (More Offspring) باقی میگذارند و فراوانی اللهای مفید (Beneficial Alleles) در جمعیت افزایش مییابد. نکتهٔ مهم این است که انتخاب بر فنوتیپ (Phenotype) – یعنی صفات قابل مشاهده – عمل میکند، در حالی که تکامل در سطح فراوانی اللها (Allele Frequencies) رخ میدهد.
نمونههای بیشماری قدرت انتخاب طبیعی را نشان میدهند:
-
ملانیسم صنعتی در شبپره فلفلی (Industrial Melanism in the Peppered Moth) نشان میدهد که چگونه یک جهش منفرد که رنگ بال را تیره کرد در محیط دودهگرفتهٔ انگلستان قرن نوزدهم به مزیت بدل شد.
-
تکامل سریع مقاومت آنتیبیوتیکی در باکتریها (Rapid Evolution of Antibiotic Resistance in Bacteria) نشان میدهد که انتخاب میتواند در بازههای زمانی بسیار کوتاه تحت فشارهای شدید محیطی رخ دهد.
این موارد نشان میدهد که انتخاب طبیعی فرایندی پیشبینانه (Forward-Looking) نیست، بلکه واکنشی پیوسته به شرایط کنونی اکولوژیک (Continual Response to Current Ecological Conditions) است.
۲.۳ رانش ژنتیکی: تکامل تصادفی (Genetic Drift: Evolution by Chance)
در حالی که انتخاب طبیعی نیرویی قطعی (Deterministic Force) است، تکامل همچنین از طریق رانش ژنتیکی (Genetic Drift) پیش میرود؛ یعنی نوسان تصادفی فراوانی اللها از نسلی به نسل دیگر. رانش در جمعیتهای کوچک اهمیت بیشتری دارد، جایی که رویدادهای شانسی – مانند اینکه کدام افراد تولیدمثل کنند – میتوانند باعث تثبیت یا حذف اللها (Fixation or Loss of Alleles) شوند، بیآنکه اثر آن اللها بر برازندگی (Fitness) مهم باشد.
دو سناریوی کلاسیک رانش عبارتاند از:
-
اثر بنیانگذار (Founder Effect): زمانی که تعداد کمی فرد زیستگاهی جدید را کلنی میکنند.
-
اثر گلوگاه (Bottleneck Effect): در پی کاهش شدید جمعیت.
در بازههای طولانی، رانش میتواند تنوع ژنتیکی (Genetic Variation) را کاهش داده و توانایی جمعیت برای سازگاری را محدود کند. حتی ممکن است جهشهای خنثی یا کمی زیانآور (Neutral or Mildly Deleterious Mutations) تثبیت شوند. تعامل بین رانش و انتخاب، موضوعی کلیدی در ژنتیک جمعیت (Population Genetics) است و قدرت نسبی هرکدام عمدتاً به اندازهٔ جمعیت (Population Size) و شدت فشارهای انتخابی بستگی دارد.
۲.۴ جریان ژن: پیوند جمعیتها (Gene Flow: Connecting Populations)
جریان ژن (Gene Flow) یا مهاجرت (Migration)، جابهجایی اللها بین جمعیتها از طریق پراکندگی افراد یا گامتها (Gametes) است. این پدیده با وارد کردن مادهٔ ژنتیکی جدید، اثر رانش و سازگاری محلی (Local Adaptation) را خنثی میکند و باعث افزایش تنوع ژنتیکی میشود.
نمونهها:
-
انتقال گرده (Pollen) توسط باد یا حشرات که اللها را بین جمعیتهای گیاهی جدا از هم جابهجا میکند.
-
مهاجرت جانوران از میان موانع جغرافیایی که الگوهای فراوانی اللها را تغییر میدهد.
جریان ژن میتواند جمعیتها را همگن (Homogenize) کند و آنها را از نظر ژنتیکی مشابهتر سازد، اما همچنین میتواند جهشهای سودمند (Advantageous Mutations) را بهسرعت در سراسر گسترهٔ یک گونه پخش کند.
۲.۵ جهش: سرچشمهٔ نهایی نوآوری (Mutation: The Ultimate Source of Novelty)
هرچند انتخاب طبیعی، رانش و جریان ژن تنوع موجود را جابهجا میکنند، اما جهش (Mutation) منبع اصلی اللهای جدید است. جهشها خودبهخود در اثر خطاهای همانندسازی DNA (Replication Errors) یا عوامل خارجی مانند اشعهٔ فرابنفش (Ultraviolet Light) و مواد جهشزا (Chemical Mutagens) رخ میدهند. این تغییرات میتوانند شامل تغییر تکنوکلئوتیدی (Single Nucleotide Changes)، درج یا حذف (Insertions/Deletions) یا بازآراییهای کروموزومی (Chromosomal Rearrangements) باشند.
بیشتر جهشها خنثی یا مضرند، ولی بخش کوچکی میتواند سودمند باشد و مواد خام نوآوری تکاملی (Raw Material for Innovation) را فراهم کند. در مقیاس زمانی تکاملی، انباشت جهشها باعث واگرایی دودمانها (Lineage Divergence) و ظهور صفات جدید میشود.
۲.۶ برازندگی، سازگاری و محیط (Fitness, Adaptation, and the Environment)
مفهوم مرکزی در تفکر تکاملی، برازندگی (Fitness) است که به موفقیت تولیدمثلی نسبی (Relative Reproductive Success) یک ارگانیسم نسبت به دیگران اشاره دارد. برازندگی به میزان تطابق صفات یک ارگانیسم با محیط آن بستگی دارد.
-
سازگاری (Adaptation) صفتی قابل وراثت (Heritable Trait) است که با بهبود بقا یا تولیدمثل، برازندگی را افزایش میدهد.
-
سازگاریها میتوانند ریختشناسی (Morphological) باشند، مانند بدن هیدرودینامیک دلفین برای شنا،
فیزیولوژیک (Physiological) مانند پروتئینهای ضدیخ در ماهیهای قطب جنوب،
یا رفتاری (Behavioral) مانند شکار گروهی در گرگها.
از آنجا که محیطها تغییر میکنند، سازگاریها وابسته به شرایط (Context-Dependent) هستند؛ صفتی که در یک محیط سودمند است ممکن است در محیطی دیگر خنثی یا حتی زیانآور شود.
۲.۷ ادغام سازوکارها: سنتز مدرن (Integrating Mechanisms: The Modern Synthesis)
اوایل قرن بیستم، سنتز مدرن (Modern Synthesis) نظریهٔ انتخاب طبیعی داروین (Darwin’s Natural Selection) را با ژنتیک مندل (Mendelian Genetics) و ژنتیک جمعیت (Population Genetics) ترکیب کرد. پیشگامانی چون رونالد فیشر (Ronald Fisher)، ج. ب. اس. هالدین (J. B. S. Haldane) و سیوال رایت (Sewall Wright) مدلهای ریاضی توسعه دادند تا نشان دهند چگونه جهش (Mutation)، انتخاب (Selection)، رانش (Drift) و مهاجرت (Migration) با هم تعامل میکنند و فراوانی اللها را در طول زمان شکل میدهند.
این ادغام نشان داد که تکامل اساساً تغییر در ترکیب ژنتیکی جمعیتها (Change in Genetic Composition of Populations) است و حتی تغییرات کوچک و پیوسته در فراوانی اللها میتواند در طول زمان به دگرگونیهای تکاملی چشمگیر (Significant Evolutionary Transformations) منجر شود.
همچنین سنتز مدرن تأکید کرد که فرآیندهای ریزتکاملی (Microevolutionary Processes) – مانند تغییر در فراوانی ژنها – برای توضیح الگوهای کلانتکاملی (Macroevolutionary Patterns) از جمله منشأ گونههای جدید و تغییرات وسیع در سابقهٔ فسیلی (Fossil Record) کافی هستند.
۲.۸ فراتر از سنتز مدرن: دیدگاههای نو (Beyond the Modern Synthesis: New Perspectives)
اگرچه سنتز مدرن پایهٔ زیستشناسی تکاملی است، پژوهشهای بعدی دامنهٔ آن را گسترش داده است:
-
نظریهٔ خنثی (Neutral Theory) که توسط موتو کیمورا (Motoo Kimura) مطرح شد، بر اهمیت جهشهای نه سودمند و نه زیانآور تأکید دارد و نشان میدهد که بخش بزرگی از تکامل مولکولی به وسیلهٔ رانش (Drift) و نه انتخاب هدایت میشود.
-
کشف انتقال افقی ژن (Horizontal Gene Transfer) – تبادل مادهٔ ژنتیکی بین موجودات نامرتبط – الگوی پیچیدهتر و شبکهایتری از تکامل را آشکار کرده است، بهویژه در میکروبها.
-
پیشرفتهای زیستشناسی تکوینی تکاملی (Evo-Devo: Evolutionary Developmental Biology) نشان داده که تغییر در تنظیم بیان ژن (Gene Regulation) میتواند نوآوریهای ریختی عمده ایجاد کند بدون آنکه نیاز به تغییرات وسیع در توالیهای کدکنندهٔ پروتئین باشد.
-
همچنین تغییرات اپیژنتیک (Epigenetic Modifications)، روابط همزیستی (Symbiotic Relationships) و نظریهٔ ساخت طاقچه (Niche Construction Theory) تعامل پیچیدهٔ ارگانیسمها با محیط را نشان میدهند.
۲.۹ جمعبندی اصول اساسی (Synthesis of Core Principles)
در مجموع، تنوع (Variation)، وراثت (Heredity)، انتخاب طبیعی (Natural Selection)، رانش ژنتیکی (Genetic Drift)، جریان ژن (Gene Flow) و جهش (Mutation) هستهٔ فرآیندهایی را تشکیل میدهند که تکامل را هدایت میکنند. این سازوکارها جداگانه عمل نمیکنند، بلکه به صورت پویا (Dynamic) و وابسته به اندازهٔ جمعیت (Population Size)، زمینهٔ بومشناختی (Ecological Context) و رخدادهای تاریخی (Historical Contingencies) با یکدیگر تعامل دارند.
به عنوان نمونه، یک جهش سودمند ممکن است در یک جمعیت بزرگ تحت انتخاب قوی به سرعت گسترش یابد، اما همان جهش میتواند در یک گروه کوچک و منزوی به دلیل شانس از بین برود. درک این تعاملات، پایهٔ مفهومی همهٔ موضوعات بعدی در زیستشناسی تکاملی است؛ از خاستگاه گونههای جدید (Origin of New Species) گرفته تا الگوهای پیچیدهٔ تنوع زیستی (Biodiversity) در سراسر سیاره.
۳. شواهد تکامل (Evidence for Evolution)
تکامل (Evolution) یک مفهوم صرفاً نظری نیست؛ بلکه توسط بدنهای گسترده و متنوع از شواهد تجربی پشتیبانی میشود که از شاخههای مختلف زیستشناسی به دست آمده است. دانشمندان تغییرات تکاملی را بهطور مستقیم در مطالعات آزمایشگاهی و میدانی مشاهده کردهاند و سوابق تاریخی موجود در فسیلها (Fossils)، آناتومی (Anatomy) و ژنومها (Genomes) این مشاهدات را تأیید میکنند. این شواهد نشان میدهند که فرآیندهای تکاملی جهانی هستند و همزمان یگانگی و تنوع حیات در زمین را آشکار میسازند.
۳.۱ سوابق فسیلی (The Fossil Record)
یکی از قویترین منابع شواهد برای تکامل، سوابق فسیلی (Fossil Record) هستند که ظهور و انقراض گونهها در طول زمان زمینشناسی را مستندسازی میکنند. فسیلها، که بقایا یا اثرات حفظشده جانداران باستانی هستند، تصویری از حیات در دورههای مختلف ارائه میدهند و به پژوهشگران امکان میدهند تا تاریخچه حیات (History of Life) را بازسازی کنند.
شکلهای انتقالی (Transitional Forms) که ویژگیهای میانه بین گروههای اجدادی و مشتق را دارند، بهویژه آموزنده هستند. برای مثال، فسیل Tiktaalik roseae با ترکیبی از ویژگیهای ماهی و چهاراندام، انتقال از زندگی آبزی به زمینی را نشان میدهد. به همین ترتیب، تکامل پرندگان از دایناسورهای تراپود (Theropod Dinosaurs) با فسیلهایی که تغییرات اسکلت، پرها و سازگاریهای دیگر را طی میلیونها سال نشان میدهند، تأیید میشود. شواهد فسیلی نشاندهنده تغییرات تدریجی در شاخهها (Lineages) و همچنین الگوهای انشعابی مشخص برای رخدادهای فرگشت گونهای (Speciation Events) هستند.
با اینکه سوابق فسیلی بهطور ذاتی ناقص هستند، به دلیل نادر بودن فسیلسازی و فرآیندهای زمینشناسی که بقایا را تخریب میکنند، الگوهای مشاهدهشده با پیشبینیهای نظریه تکامل هماهنگاند و ارزش شواهدی آنها را تقویت میکنند. روشهای لایهبندی زمینشناسی (Stratigraphy)، تاریخگذاری رادیومتریک (Radiometric Dating) و آناتومی مقایسهای (Comparative Morphology) همگی چارچوب زمانی فراهم میکنند که در آن میتوان فرآیندهای تکاملی را مطالعه و کمّیسازی کرد.
۳.۲ آناتومی مقایسهای (Comparative Anatomy)
آناتومی مقایسهای (Comparative Anatomy) شواهد بیشتری ارائه میدهد و شباهتهای ساختاری بین جانداران را نشان میدهد که منعکسکننده تبار مشترک (Common Ancestry) است.
ساختارهای همریشه (Homologous Structures) مانند اندامهای جلویی (Forelimbs) در پستانداران، پرندگان و خزندگان، ساختار استخوانی مشابهی دارند هرچند برای عملکردهای متفاوتی مانند گرفتن، پرواز یا شنا سازگار شدهاند. این شباهتها نشان میدهند که این جانداران از جد مشترک (Common Ancestor) برخوردار بودهاند که اندام جلویی عمومی داشته است.
در مقابل، ساختارهای مشابه (Analogous Structures) مانند بالهای پرندگان و حشرات، بهطور مستقل در نتیجه فرگشت همگرا (Convergent Evolution) پدید آمدهاند و نشان میدهند که فشارهای انتخابی مشابه میتوانند سازگاریهای مشابه در گونههای نامرتبط ایجاد کنند.
ساختارهای باقیمانده (Vestigial Structures) مانند آپاندیس انسان، استخوانهای لگنی در نهنگها و بالهای ناقص در پرندگان پروازنکردنی، نیز تاریخچه تکاملی را نشان میدهند، زیرا این صفات در اجداد کارکرد داشتهاند اما اکنون ضروری نیستند.
آناتومی مقایسهای فراتر از اسکلت است. ساختارهای بافت نرم، سیستمهای اندامی و الگوهای تکاملی (Developmental Patterns) نیز ویژگیهای حفظشدهای را نشان میدهند که به اجداد مشترک قدیمی برمیگردند. برای مثال، چیدمان قوسهای حلقی (Pharyngeal Arches) در جنینهای مهرهداران، مسیرهای توسعه مشترک را نشان میدهد و همریختی عمیق (Deep Homology) موجود در اشکال بزرگسال متفاوت را آشکار میکند.
۳.۳ جنینشناسی و رشد (Embryology and Development)
جنینشناسی (Embryology) با مقایسه فرآیندهای توسعهای بین گونهها، روابط تکاملی را روشن میکند. مراحل ابتدایی جنینهای مهرهداران شباهتهای چشمگیری نشان میدهند، مانند کیسههای حلقی (Pharyngeal Pouches)، سومیتها (Somites) و دم (Tail Structures) که بازتاب تبار مشترک هستند.
این ویژگیهای جنینی معمولاً پیش از ظاهر شدن ساختارهای بزرگسال مربوطه وجود دارند و نشان میدهند که تغییرات تکاملی در توسعه میتوانند مورفولوژیهای جدید بزرگسال را بدون تغییر الگوهای ابتدایی جنینی ایجاد کنند.
زیستشناسی توسعهای تکاملی (Evolutionary Developmental Biology یا Evo-Devo) این دیدگاه را گسترش داده است و ژنها، شبکههای تنظیمی و مسیرهای سیگنالدهی (Signaling Pathways) را شناسایی کرده که توسعه را کنترل میکنند. حفظ ژنهای کلیدی توسعهای (مثل خوشههای ژنی Hox) در شاخههای متنوع، شواهد مولکولی تبار مشترک را ارائه میدهد و نشان میدهد چگونه تغییرات کوچک در بیان ژنها میتوانند نوآوریهای مورفولوژیکی گسترده ایجاد کنند.
۳.۴ شواهد مولکولی و ژنومی (Molecular and Genomic Evidence)
عصر مولکولی (Molecular Era) شواهد قدرتمندی برای تکامل ارائه کرده است از طریق مقایسه توالی DNA، RNA و پروتئینها. جاندارانی که جد مشترک نزدیک دارند، شباهت بیشتری در توالی ژنتیکی خود دارند، در حالی که گونههای دورتر با گذشت زمان تفاوتها را انباشته میکنند.
برای مثال، انسانها تقریباً ۹۸–۹۹٪ DNA خود را با شامپانزهها به اشتراک میگذارند که نشاندهنده رابطه تکاملی نزدیک آنها است. درختان فیلوژنتیک (Phylogenetic Trees) که از دادههای مولکولی استخراج شدهاند، رابطهها را مطابق با آنچه از مورفولوژی و سوابق فسیلی نتیجهگیری شده است نشان میدهند و همخوانی نظریه تکامل در شواهد مختلف را تأیید میکنند.
شواهد مولکولی همچنین ژنها، پروتئینها و مسیرهای بیوشیمیایی حفظشده را نشان میدهند که تقریباً در تمام موجودات زندگی مشترکاند. کد ژنتیکی (Genetic Code) تقریباً در باکتریها، گیاهان و حیوانات یکسان است و نشان میدهد که تمام موجودات از یک جد مشترک مشتق شدهاند. ژنهای غیر فعال یا ژنهای شبه (Pseudogenes) نیز شواهدی اضافی ارائه میدهند، زیرا در مکانهای متناظر در گونههای مرتبط یافت میشوند و الگوهای جهش آنها با تبار مشترک همخوانی دارد.
۳.۵ جغرافیای زیستی (Biogeography)
جغرافیای زیستی (Biogeography)، مطالعه توزیع جغرافیایی گونهها، آزمایشهای طبیعی (Natural Experiments) را برای روشن کردن تاریخچه تکاملی فراهم میکند. الگوهای توزیع گونهها اغلب با رویدادهای تاریخی مانند رانش قارهای (Continental Drift)، ایزولاسیون در جزایر و قطعهبندی زیستگاهها همخوانی دارد.
برای مثال، جانوران منحصربهفرد استرالیا، از جمله پستانداران کیسهای مانند کانگورو و وومبتها (Wombats)، عمدتاً در ایزولاسیون تکامل یافتهاند پس از جدا شدن قاره از گوندوانا (Gondwana). همچنین، فنچهای داروین (Darwin’s Finches) در جزایر گالاپاگوس نمونهای از تابعگستری سازگار (Adaptive Radiation) هستند، که در آن یک گونه اجدادی به گونههای متعدد در هر زیستگاه تخصصی تبدیل شده است. این الگوها بدون فرگشت توضیح داده نمیشوند و نشان میدهند که جغرافیا و فشارهای محیطی، تنوع را شکل میدهند.
۳.۶ شواهد تجربی و مشاهدهای (Experimental and Observational Evidence)
تکامل تنها یک فرآیند تاریخی نیست؛ بلکه میتوان آن را در زمان واقعی نیز مشاهده کرد. میکروارگانیسمها، با زمان تولید مثل کوتاه، نمونههای برجستهای از تکامل در عمل ارائه میدهند.
برای مثال، آزمایشها با Escherichia coli در محیطهای کنترلشده ظهور تواناییهای متابولیک جدید، مقاومت به آنتیبیوتیکها و مسیرهای تکاملی قابل پیشبینی را طی هزاران نسل نشان دادهاند. در طبیعت، جمعیتهای حشرات تحت فشار آفتکشها طی چند سال مقاومت ایجاد میکنند و نشاندهنده انتخاب طبیعی (Natural Selection) در عمل هستند. تغییرات فصلی در فراوانی آللها در جمعیتهای وحشی، مانند رنگ بدن در پروانهها یا ضخامت پوشش بدن در جوندگان، شواهد مستقیم دیگری از دینامیک تکاملی در محیط طبیعی ارائه میدهند.
۳.۷ همگرایی چندین خط شواهد (Integrating Multiple Lines of Evidence)
قدرت شواهد تکامل نه در یک منبع منفرد، بلکه در همگرایی چندین منبع مستقل نهفته است. فسیلها، آناتومی مقایسهای، جنینشناسی، بیولوژی مولکولی، جغرافیای زیستی و مشاهده مستقیم همگی داستانی سازگار را روایت میکنند: زندگی روی زمین از طریق تبار مشترک به هم متصل است و جمعیتها تحت تأثیر انتخاب طبیعی، رانش ژنتیکی، جریان ژنی و جهش تغییر میکنند.
هماهنگی و پیشبینیپذیری این الگوها پایه علمی محکمی فراهم میکند که بیش از ۱۶۰ سال تحت بررسی دقیق قرار داشته است و همچنان راهنمای پژوهش در ژنتیک، اکولوژی، پزشکی و زیستشناسی تکاملی است.
۴. مکانیسمهای تغییر تکاملی (Mechanisms of Evolutionary Change)
تغییر تکاملی (Evolutionary Change) از طریق چندین فرآیند بههمپیوسته رخ میدهد که ترکیب ژنتیکی (Genetic Composition) و صفات فنوتیپی (Phenotypic Traits) جمعیتها را در طول زمان تغییر میدهند. درک این مکانیسمها برای توضیح سازگاریهای کوچک درون جمعیتها و همچنین الگوهای بزرگمقیاس مشاهدهشده در تاریخ حیات ضروری است. اگرچه انتخاب طبیعی (Natural Selection)، رانش ژنتیکی (Genetic Drift)، جریان ژنی (Gene Flow) و جهش (Mutation) هسته اصلی این فرآیندها را تشکیل میدهند، مکانیسمهای دیگری مانند انتخاب جنسی (Sexual Selection)، همتکامل (Coevolution) و تعاملات بومشناختی (Ecological Interactions) نیز به تنوع و پیچیدگی نتایج تکاملی کمک میکنند.
۴.۱ ریزتکامل در مقابل فرگشت کلان (Microevolution versus Macroevolution)
تکامل در مقیاسهای مختلف (Multiple Scales) عمل میکند.
ریزتکامل (Microevolution) به تغییرات فرکانس آللها (Allele Frequencies) و توزیع صفات (Trait Distributions) در جمعیتها طی بازههای زمانی نسبتاً کوتاه اشاره دارد. نمونهها شامل ایجاد مقاومت به آنتیبیوتیک در باکتریها، تغییرات فصلی در رنگ پوشش بدن جوندگان و تغییر زمان گلدهی گیاهان هستند. تغییرات ریزتکاملی را میتوان بهطور مستقیم در مطالعات آزمایشگاهی و میدانی مشاهده کرد و آنها شواهد ملموس دینامیک تکاملی را ارائه میدهند.
ریزتکامل همچنین پایه و اساس فرگشت کلان (Macroevolution) را فراهم میکند، که الگوهای تکاملی بزرگمقیاس را شامل میشود، مانند ایجاد گونههای جدید (Origin of New Species)، تنوعبخشی گروههای اصلی تاکسونومیک (Diversification of Major Taxonomic Groups) و ظهور صفات مورفولوژیکی و فیزیولوژیکی جدید. الگوهای فرگشت کلان معمولاً از سوابق فسیلی، آناتومی مقایسهای و درختهای فیلوژنتیک مولکولی استنتاج میشوند، اما در نهایت ریشه در تأثیرات تجمعی فرآیندهای ریزتکاملی طی دورههای طولانی دارند.
۴.۲ گونهزایی: منشأ گونههای جدید (Speciation: The Origin of New Species)
گونهزایی (Speciation) فرآیندی است که طی آن جمعیتها تفاوت پیدا کرده و گونههای متمایز (Distinct Species) ایجاد میکنند. این فرآیند معمولاً زمانی رخ میدهد که جمعیتها از نظر تولیدمثلی جدا شوند (Reproductively Isolated) و جریان ژنی بین آنها متوقف شود.
چندین نوع گونهزایی وجود دارد:
-
گونهزایی جداافتاده (Allopatric Speciation): رایجترین نوع است و زمانی رخ میدهد که جمعیتها بهطور جغرافیایی توسط موانعی مانند کوهها، رودخانهها یا جزایر اقیانوسی جدا شوند. این جدایی امکان تکامل مستقل را فراهم میکند و اغلب منجر به تجمع تفاوتهای ژنتیکی میشود که مانع از تولیدمثل متقابل در صورت برخورد مجدد جمعیتها میگردد.
-
گونهزایی هممحدودهای (Sympatric Speciation): در محدودههای جغرافیایی همپوشان رخ میدهد، معمولاً ناشی از تخصص اکولوژیکی، تفاوتهای رفتاری یا پلیپلوئیدی (Polyploidy) در گیاهان است.
-
گونهزایی پاراپاتریک (Parapatric Speciation): حالت میانی است، جایی که جمعیتهای هممرز در حالی که جریان ژن محدود دارند، تفاوت پیدا میکنند، اغلب در طول گرادیانهای محیطی.
-
مناطق هیبریدی (Hybrid Zones): مناطقی که در آن جمعیتهای نیمه جداشده تولیدمثلی با هم برخورد میکنند، تجمع تدریجی موانع تولیدمثلی و تعامل پیچیده انتخاب و جریان ژن را نشان میدهند.
گونهزایی میتواند سریع یا کند باشد، بسته به فشارهای محیطی، اندازه جمعیت و معماری ژنتیکی. مثالهایی مانند تنوع فنچهای داروین یا ماهیان سیکلید در دریاچههای آفریقا نشان میدهند که گونههای جدید میتوانند به سرعت در شرایط مساعد و فشارهای انتخابی مناسب ایجاد شوند.
۴.۳ انتخاب جنسی و استراتژیهای تولیدمثلی (Sexual Selection and Reproductive Strategies)
انتخاب جنسی (Sexual Selection) نوعی انتخاب طبیعی ویژه است که ناشی از تفاوت در موفقیت تولیدمثلی (Reproductive Success) به دلیل رقابت برای جفتها (Competition for Mates) است. صفاتی که جذابیت فرد را افزایش داده یا موفقیت در رقابت درونجنسی را بهبود میبخشند، حتی اگر هزینههای بقا داشته باشند، ممکن است تکامل یابند.
نمونههای کلاسیک شامل:
-
پرهای پر زرق و برق طاووس نر
-
شاخهای بزرگ گوزنها
-
نمایشهای جفتگیری بسیاری از پرندگان و ماهیان
انتخاب جنسی میتواند دیمورفیسم جنسی (Sexual Dimorphism) چشمگیری ایجاد کند و تکامل صفات رفتاری، مورفولوژیکی و فیزیولوژیکی را هدایت نماید. گاهی انتخاب جنسی با انتخاب طبیعی تعامل دارد و منجر به تضاد بین موفقیت جفتیابی و بقا (Trade-offs) میشود که مسیرهای تکاملی را پیچیده میکند.
۴.۴ انتخاب خویشاوندی و رفتارهای نوعدوستانه (Kin Selection and Altruistic Behavior)
تمام تغییرات تکاملی ناشی از بقا و موفقیت فردی نیستند. انتخاب خویشاوندی (Kin Selection) توضیح میدهد که چگونه رفتارهایی که کاهشدهنده موفقیت مستقیم فرد (Direct Fitness) هستند، میتوانند تکامل یابند اگر موفقیت تولیدمثلی خویشاوندان حامل ژنهای مشابه افزایش یابد.
رفتارهای نوعدوستانه (Altruistic Behaviors) مانند:
-
تولیدمثل مشارکتی در پرندگان
-
اعلان خطر در سنجابهای زمینی (Ground Squirrels)
-
تقسیم غذا در حشرات اجتماعی
نگهداری میشوند زیرا بقا و تولیدمثل خویشاوندان ژنتیکی را افزایش میدهند. مفهوم بقای جامع (Inclusive Fitness) که هم موفقیت تولیدمثلی مستقیم و هم غیرمستقیم را در نظر میگیرد، چارچوبی برای درک این الگوها فراهم میکند و دامنه مکانیسمهای تکاملی را فراتر از انتخاب سطح فردی توسعه میدهد.
۴.۵ همتکامل و تعاملات بومشناختی (Coevolution and Ecological Interactions)
همتکامل (Coevolution) زمانی رخ میدهد که دو یا چند گونه تکامل یکدیگر را بهطور متقابل تحت تأثیر قرار دهند. نمونههای کلاسیک شامل:
-
دینامیک شکار و شکارچی (Predator-Prey Dynamics)
-
تعامل میزبان و انگل (Host-Parasite Interactions)
-
روابط همزیستی مانند گردهافشانها و گیاهان گلدار (Mutualistic Relationships)
در این سناریوها، سازگاریها در یک گونه فشار انتخابی بر دیگری اعمال میکنند و منجر به مسابقه تسلیحاتی تکاملی (Evolutionary Arms Races) یا ویژگیهای متقابلاً یکپارچه همزیستی میشوند. برای مثال، نیشهای بلند برخی ارکیدهها با خرطوم حشرات گردهافشان خاص همتکامل یافتهاند، که انتخاب متقابل طی بازههای تکاملی طولانی را نشان میدهد.
همتکامل وابستگی جانداران در اکوسیستمها را روشن میکند و نشان میدهد که مکانیسمهای تکاملی نه بهطور جداگانه، بلکه بهعنوان بخشی از شبکههای پیچیده بومشناختی عمل میکنند.
۴.۶ رانش ژنتیکی، جریان ژن و جهش در زمینه مکانیسمی (Genetic Drift, Gene Flow, and Mutation in Mechanistic Context)
در حالی که انتخاب و همتکامل تغییرات سازگارانه را شکل میدهند، رانش ژنتیکی (Genetic Drift)، جریان ژن (Gene Flow) و جهش (Mutation) مکانیسمهای دیگری برای تغییر تکاملی فراهم میکنند.
-
رانش ژنتیکی (Drift): نوسانات تصادفی در فرکانس آللها ایجاد میکند و میتواند منجر به ثابت شدن یا از بین رفتن تصادفی آللها شود، بهویژه در جمعیتهای کوچک.
-
جریان ژن (Gene Flow): جمعیتها را به هم متصل میکند، امکان انتشار آللهای مفید را فراهم میکند و از تفاوت بیش از حد جمعیتها جلوگیری مینماید.
-
جهش (Mutation): متغیرهای ژنتیکی جدید (New Genetic Variants) ایجاد میکند که پتانسیل سازگاری (Adaptation Potential) را فراهم میسازد.
ترکیب این مکانیسمها نتایج تکاملی متغیر و وابسته به زمینه (Context-Dependent) تولید میکند و تعامل بین تصادف، فشارهای محیطی و شرایط تاریخی را نشان میدهد.
۴.۷ چشماندازهای سازگاری و مسیرهای تکاملی (Adaptive Landscapes and Evolutionary Trajectories)
مفهوم چشماندازهای سازگاری (Adaptive Landscapes) که توسط سوالال رایت (Sewall Wright) ارائه شد، چارچوب مفهومی برای درک چگونگی تکامل جمعیتها در طول چشماندازهای بقا (Fitness Landscapes) ارائه میکند.
-
قلهها (Peaks): ترکیبهای با بقای بالا (High-Fitness Combinations)
-
درهها (Valleys): ترکیبهای با بقای پایین (Low-Fitness Combinations)
جمعیتها میتوانند از طریق جهش، بازترکیب (Recombination) و انتخاب در این چشماندازها حرکت کنند و به قلههای محلی یا جهانی (Local or Global Fitness Peaks) برسند. این استعاره توضیح میدهد که چرا تکامل ممکن است فرمهای بسیار سازگار ایجاد کند که با این حال توسط محدودیتهای تاریخی، همبستگیهای ژنتیکی و محدودیتهای محیطی محدود شدهاند. مسیرهای تکاملی (Evolutionary Trajectories) تحت تأثیر محیط انتخابی فعلی و معماری ژنتیکی جمعیتها شکل میگیرند.
۴.۸ ادغام مکانیسمها در جمعیتهای طبیعی (Integration of Mechanisms in Natural Populations)
در طبیعت، مکانیسمهای تکاملی بهندرت بهطور جداگانه عمل میکنند. انتخاب، رانش ژنتیکی، جریان ژن، جهش، انتخاب جنسی و همتکامل همزمان عمل کرده و سازگاری کوتاهمدت و تنوع بلندمدت را شکل میدهند.
درک این تعاملها برای توضیح الگوهای تغییر، گونهزایی و انقراض در سراسر درخت حیات (Tree of Life) ضروری است. زیستشناسی تکاملی مدرن از مشاهدات میدانی، آزمایشهای آزمایشگاهی و مدلسازی محاسباتی (Computational Modeling) استفاده میکند تا این دینامیکها را تحلیل و چارچوبهای پیشبینیکننده برای پاسخهای تکاملی به تغییرات محیطی، فعالیت انسانی و بیماریهای نوظهور ارائه دهد.
۵. دیدگاههای مولکولی و ژنتیکی (Molecular and Genetic Perspectives)
درک مدرن تکامل (Evolution) بهطور عمیق در زیستشناسی مولکولی (Molecular Biology) و ژنتیک (Genetics) ریشه دارد، که پایهای مکانیکی برای وراثت، تغییر و اصلاح صفات (Inheritance, Variation, and Modification of Traits) در نسلها فراهم میکنند. در حالی که نظریه کلاسیک تکامل عمدتاً بر صفات فنوتیپی (Phenotypic Traits) قابل مشاهده در جمعیتها تمرکز داشت، پیشرفتهای زیستشناسی مولکولی به دانشمندان امکان میدهد تغییرات ژنتیکی اساسی که راننده تکامل هستند را بررسی کنند. ژنها (Genes)، ژنومها (Genomes) و مسیرهای مولکولی (Molecular Pathways) زمینهای هستند که نیروهای تکاملی بر آن عمل میکنند و ریزتکامل (Microevolution) را به فرگشت کلان (Macroevolution) متصل مینمایند.
۵.۱ ژنتیک جمعیت: کمیسازی تکامل (Population Genetics: Quantifying Evolution)
ژنتیک جمعیت (Population Genetics) چارچوب ریاضی است که توضیح میدهد چگونه فرکانس آللها (Allele Frequencies) در طول زمان در جمعیتها تغییر میکند. این رشته مفاهیم جهش (Mutation)، انتخاب (Selection)، رانش ژنتیکی (Genetic Drift) و جریان ژن (Gene Flow) را در مدلهای پیشبینیکننده دینامیک تکاملی (Evolutionary Dynamics) ادغام میکند.
کارهای بنیادی رونالد فیشر (Ronald Fisher)، جی. بی. اس. هالدین (J. B. S. Haldane) و سوال رایت (Sewall Wright) روابط بین این فرآیندها و پارامترهای جمعیت مانند اندازه جمعیت (Population Size)، ساختار جمعیت (Population Structure) و الگوهای جفتگیری (Mating Patterns) را به صورت رسمی بیان کردند. ژنتیک جمعیت به زیستشناسان امکان میدهد قدرت انتخاب طبیعی (Strength of Natural Selection) را اندازهگیری، احتمال ثابت شدن آللها (Allele Fixation) را پیشبینی و نقش رویدادهای تصادفی در شکلدهی تنوع ژنتیکی (Genetic Variation) را ارزیابی کنند.
ژنتیک جمعیت همچنین پایهای برای درک پدیدههای پیچیدهتر مانند:
-
مزیت هتروزیگوت (Heterozygote Advantage)
-
انتخاب وابسته به فرکانس (Frequency-Dependent Selection)
-
حفظ پلیمورفیسمها (Maintenance of Polymorphisms)
با استفاده از اصول ژنتیک جمعیت، پژوهشگران میتوانند دادههای تجربی DNA و توالی پروتئینها را تجزیه و تحلیل کرده، تاریخچه تکاملی (Evolutionary History) را بازسازی، زمانهای انشعاب (Divergence Times) را تخمین و نقش نیروهای تکاملی مختلف را ارزیابی کنند. بنابراین ژنتیک جمعیت شکاف بین تغییرات قابل مشاهده و مکانیزمهای مولکولی تکامل را پر میکند.
۵.۲ نظریه خنثی و ساعت مولکولی (Neutral Theory and Molecular Clocks)
یکی از پیشرفتهای بزرگ در تکامل مولکولی نظریه خنثی (Neutral Theory) است که توسط موتو کیمورا (Motoo Kimura) ارائه شد. این نظریه بیان میکند که اکثر جهشهای ژنتیکی در سطح مولکولی خنثی هستند و عمدتاً توسط رانش ژنتیکی (Genetic Drift) تثبیت یا از بین میروند، نه انتخاب طبیعی (Natural Selection).
نظریه خنثی چارچوب قدرتمندی برای تفسیر الگوهای تنوع مولکولی (Patterns of Molecular Variation) فراهم میکند و توضیح میدهد چرا برخی نواحی ژنوم کندتر از سایر بخشها تکامل مییابند. جهشهای هممعنی (Synonymous Mutations) که توالی آمینواسید را تغییر نمیدهند، معمولاً با نرخ قابل پیشبینی انباشته میشوند و پایهای برای مفهوم ساعت مولکولی (Molecular Clock) ایجاد میکنند.
ساعت مولکولی به زیستشناسان امکان میدهد زمان انشعابات تکاملی (Evolutionary Divergences) را با مقایسه تفاوتهای توالی بین گونهها تخمین بزنند. با کالیبراسیون ساعت با دادههای فسیلی یا زمینشناسی (Fossil or Geological Data)، پژوهشگران میتوانند زمانبندی گونهزایی (Speciation Events)، الگوهای مهاجرت (Migration Patterns) و ظهور صفات کلیدی را بازسازی کنند. ساعت مولکولی در تاکسونهای مختلف از میکروبها تا نخستیسانان به کار رفته و درک ما از جدول زمانی تکامل (Evolutionary Timelines) را متحول کرده است.
۵.۳ انتقال ژن افقی و تکامل شبکهای (Horizontal Gene Transfer and Reticulate Evolution)
در حالی که وراثت عمودی (Vertical Inheritance) از والد به فرزند غالب است، انتقال ژن افقی (Horizontal Gene Transfer, HGT) نقش مهمی در تکامل بسیاری از جانداران، بهویژه پروکاریوتها (Prokaryotes) ایفا میکند. HGT اجازه میدهد ژنها بین گونههای نامرتبط منتقل شوند و باعث کسب سریع صفات جدید مانند مقاومت به آنتیبیوتیک یا تواناییهای متابولیکی گردد.
این فرآیند مرزهای سنتی شجرهای (Lineage Boundaries) را مبهم کرده و الگوهای تکاملی شبکهای (Reticulate Evolutionary Patterns) ایجاد میکند، بهویژه در جوامع میکروبی. HGT نشان میدهد که تکامل همیشه درختی نیست (Tree-Like)؛ بلکه میتواند شامل شبکههای تبادل ژنتیکی باشد که بازسازی فیلوژنتیکی را پیچیده کرده و همزمان سازگاری را تسریع میکنند.
۵.۴ مکانیزمهای مولکولی تنوع (Molecular Mechanisms of Variation)
در سطح مولکولی، تکامل توسط ترکیبی از:
-
جهش نقطهای (Point Mutations)
-
درجها و حذفها (Insertions and Deletions)
-
تکثیر ژنها (Gene Duplications)
-
واژگونیها (Inversions)
-
بازچینیهای کروموزومی (Chromosomal Rearrangements)
رهبری میشود. به ویژه تکثیر ژن (Gene Duplications) ماده خام برای نوآوری فراهم میکند، به گونهای که یک نسخه وظایف ضروری را حفظ کرده و نسخه دیگر جهشهایی کسب میکند که میتواند صفات یا عملکردهای جدید ایجاد کند.
عناصر متحرک ژنتیکی (Transposable Elements)، توالیهای تکراری (Repetitive Sequences) و عناصر ژنتیکی موبایل (Mobile Genetic Elements) به بازسازی ژنوم و تکامل تنظیمی (Regulatory Evolution) کمک میکنند. تغییرات اپیژنتیکی (Epigenetic Modifications)، از جمله متیلاسیون DNA و تغییرات هیستونی (Histone Modification)، میتوانند الگوهای بیان ژن و نتایج فنوتیپی را تحت تأثیر قرار دهند و مسیرهای تکاملی را حتی در غیاب تغییرات توالی DNA شکل دهند.
۵.۵ فیلوژنومیک و ژنومیک تطبیقی (Phylogenomics and Comparative Genomics)
ظهور توالییابی کل ژنوم (Whole-Genome Sequencing) امکان فیلوژنومیک (Phylogenomics) را فراهم کرد، که نظریه تکامل را با دادههای جامع ژنتیکی برای بازسازی روابط گونهها و شاخهها (Lineages) تلفیق میکند.
ژنومیک تطبیقی (Comparative Genomics) توالیهای محافظتشده، خانوادههای ژنی و شبکههای تنظیمی (Regulatory Networks) را در تاکسونها بررسی میکند و بینشهایی درباره اشتراکات اجدادی (Shared Ancestry) و نوآوریهای خاص خط تکاملی (Lineage-Specific Innovations) ارائه میدهد.
مثالها:
-
تحلیل ژنهای هوموباکس (Homeobox Genes) در حیوانات، برنامههای توسعهای محافظتشدهای را نشان میدهد که پایه مورفولوژیهای متنوع هستند.
-
تفاوتها در عناصر تنظیمی (Regulatory Elements) صفات خاص گونهها را توضیح میدهند.
فیلوژنومیک همچنین امکان شناسایی انتقال ژن افقی (Horizontal Gene Transfer)، نفوذ ژنتیکی (Introgression) و هیبریداسیون (Hybridization) را فراهم میکند و پیچیدگی تکامل در سطح ژنوم را نشان میدهد.
۵.۶ شواهد مولکولی برای الگوهای تکاملی (Molecular Evidence for Evolutionary Patterns)
دادههای مولکولی الگوهای مشاهدهشده در فسیلها، آناتومی و جنینشناسی (Embryology) را تأیید میکنند.
-
توالیهای همولوگ (Homologous Sequences) در گونهها نشاندهنده جد مشترک (Common Ancestry) هستند.
-
مقایسه ژنهای اورتوگ (Orthologous) و پارالوگ (Paralogous) جهشهای تکثیری ژن و واگرایی عملکردی را روشن میکند.
-
پسوژنها (Pseudogenes)، عناصر غیرکدکننده محافظتشده (Conserved Noncoding Elements) و توالیهای تکراری نشانگر تاریخچه تکاملی هستند.
DNA میتوکندریایی (Mitochondrial DNA)، DNA کلروپلاست (Chloroplast DNA) و RNA ریبوزومی (Ribosomal RNA) برای استنتاج روابط فیلوژنتیکی بسیار مفیدند، زیرا نسبتاً سریع تکامل مییابند و وراثت مادری یا اندامکی دارند. این نشانههای مولکولی بهطور جمعی تکامل جهانی را تأیید کرده و بازسازی دقیق تاریخچه خط تکاملی را ممکن میسازند.
۵.۷ ادغام دیدگاههای مولکولی و کلاسیک (Integrating Molecular and Classical Perspectives)
دیدگاههای مولکولی و ژنتیکی جایگزین شواهد کلاسیک نیستند، بلکه آنها را تکمیل میکنند و پایه مکانیکی برای درک تکامل فراهم میآورند.
-
سوابق فسیلی (Fossil Record): نتایج تاریخی فرآیندهای تکاملی را نشان میدهد.
-
آناتومی مقایسهای و جنینشناسی: الگوهای توسعهای مشترک را نمایان میکنند.
-
دادههای مولکولی: تغییرات ژنتیکی خاص که این نتایج را ایجاد میکنند، روشن میسازند.
ترکیب این رویکردها تصویر منسجمی از تکامل ارائه میدهد (Linking Genotype to Phenotype) و ریزتکامل را به فرگشت کلان متصل میکند. این دیدگاه یکپارچه به پژوهشگران امکان میدهد:
-
پیشبینی مسیرهای تکاملی آینده (Future Evolutionary Trajectories)
-
درک ظهور جهشهای بیماریزا (Disease-Causing Mutations)
-
بررسی پایه ژنتیکی سازگاری در محیطهای مختلف
۵.۸ آینده پژوهشهای تکامل مولکولی (The Future of Molecular Evolutionary Research)
پیشرفتهای توالییابی پرسرعت (High-Throughput Sequencing)، ژنومیک تکسلولی (Single-Cell Genomics) و ژنومیک کاربردی (Functional Genomics) درک ما از تکامل در سطح مولکولی را عمیقتر میکنند.
تکنولوژیهایی مانند:
-
ویرایش ژنوم مبتنی بر CRISPR (CRISPR-Based Genome Editing)
-
توالییابی بلندخوان (Long-Read Sequencing)
امکان آزمون تجربی فرضیات تکاملی (Experimental Testing of Evolutionary Hypotheses) را فراهم میکنند، از جمله بررسی پیامدهای عملکردی جهشها و تغییرات تنظیمی.
مطالعات تکامل میکروبی، تکامل ویروسها و تکامل سرطان دینامیکهای زمان واقعی تکامل مولکولی (Real-Time Molecular Evolution) را در پاسخ به فشارهای محیطی و انتخابی نشان میدهند.
با پیشرفت بیوانفورماتیک (Bioinformatics)، زیستشناسی سیستمی (Systems Biology) و زیستشناسی مصنوعی (Synthetic Biology)، دیدگاههای مولکولی رویکردهای نظری و کاربردی زیستشناسی تکاملی را شکل داده و بینش بیسابقهای درباره مکانیزمهای ایجاد و حفظ تنوع حیات فراهم میآورند.
۶. رویدادهای اصلی تکاملی (Major Evolutionary Events)
تاریخ حیات روی زمین با مجموعهای از رویدادهای تکاملی عمیق (Profound Evolutionary Events) مشخص شده است که تنوع، پیچیدگی و پراکندگی جانداران (Diversity, Complexity, and Distribution of Organisms) را شکل دادهاند. از منشاء حیات (Origin of Life) تا ظهور انسان، این نقاط عطف بازتاب فرآیندهای تدریجی و گذارهای چشمگیر (Gradual Processes and Dramatic Transitions) هستند که بایوسفر (Biosphere) را به طور بنیادی تغییر دادهاند. مطالعه این رویدادها بینش عمیقی درباره مکانیزمها، زمانبندی و پیامدهای تغییرات تکاملی فراهم میکند.
۶.۱ منشاء حیات (Origin of Life)
منشاء حیات (Origin of Life) یا ابیژنز (Abiogenesis) نشاندهنده گذار از سیستمهای شیمیایی غیرزنده (Non-Living Chemical Systems) به جانداران خودتکثیر شونده و متابولیک فعال (Self-Replicating, Metabolically Active Organisms) است.
اگرچه مسیر دقیق هنوز موضوع تحقیقات جاری است، چندین فرضیه پیشنهاد میکنند که حیات از مولکولهای آلی ساده (Simple Organic Molecules) که در محیطهای پیشزیستی (Prebiotic Environments) شکل گرفتهاند، پدید آمده است.
آزمایش میلر-یوری (Miller-Urey Experiment) نشان داد که اسیدهای آمینه (Amino Acids) و سایر ترکیبات آلی میتوانند تحت شرایطی شبیهسازیشده از زمین اولیه تشکیل شوند. این مولکولها احتمالاً به پلیمرهای پیچیدهتر (Complex Polymers) متصل شدند و در نهایت مولکولهای RNA خودتکثیر شونده (Self-Replicating RNA or RNA-Like Molecules) ایجاد شدند.
فرضیه دنیای RNA (RNA World Hypothesis) بیان میکند که مولکولهای RNA در ابتدا هم بهعنوان مواد ژنتیکی (Genetic Material) و هم بهعنوان کاتالیزور (Catalytic Entities) عمل میکردند و بعدها ژنومهای مبتنی بر DNA و آنزیمهای پروتئینی (DNA-Based Genomes and Protein Enzymes) شکل گرفتند.
منشاء حیات زمینه را برای فرآیندهای تکاملی بعدی فراهم کرد، زیرا اولین جمعیتها را با قابلیتهای جهش (Mutation)، تکثیر (Replication) و انتخاب طبیعی (Natural Selection) ایجاد نمود.
۶.۲ تکامل پروکاریوتها و یوکاریوتها (Evolution of Prokaryotes and Eukaryotes)
زندگی پروکاریوتی (Prokaryotic Life) که شامل باکتریها و آرکئاها (Bacteria and Archaea) است، نمایانگر قدیمیترین شکلهای سازمان سلولی (Earliest Forms of Cellular Organization) میباشد. این جانداران ساده تکسلولی (Simple, Unicellular Organisms) میلیاردها سال زمین را تحت سلطه داشتند و مسیرهای متابولیکی متنوعی مانند:
-
فتوسنتز (Photosynthesis)
-
ثابتسازی نیتروژن (Nitrogen Fixation)
-
شیمیسنگخواری (Chemolithotrophy)
توسعه دادند.
ظهور فتوسنتز اکسیژنزا (Oxygenic Photosynthesis) در سیانوباکتریها (Cyanobacteria) منجر به رویداد بزرگ اکسیژنزایی (Great Oxidation Event) حدود ۲.۴ میلیارد سال پیش شد و جو زمین را به طور چشمگیری تغییر داد و مسیر متابولیسم هوازی (Aerobic Metabolism) را باز کرد. این رویداد تأثیرات اکولوژیکی و تکاملی عمیقی داشت، از جمله انقراض بسیاری از جانداران بیهوازی (Anaerobic Organisms) و تنوع خطهای تکاملی هوازی (Diversification of Aerobic Lineages).
ظهور یوکاریوتها (Eukaryotes)، یعنی سلولهایی با اندامکهای غشایی (Membrane-Bound Organelles)، نقطه عطف دیگری بود. فرضیه آندوسیمبیوز (Endosymbiotic Theory) منشأ میتوکندریها و کلروپلاستها (Mitochondria and Chloroplasts) را توضیح میدهد، به این صورت که پروکاریوتهای آزاد زندگی که بعداً وارد سلولهای میزبان شدند، تبدیل به اندامک شدند.
سلولهای یوکاریوتی پیچیدگی سلولی بیشتر، ژنومهای بزرگتر و توسعه چندسلولی (Multicellularity) را ممکن کردند و زمینه را برای تنوع گیاهان، حیوانات، قارچها و پروتیستها فراهم آوردند.
۶.۳ ظهور چندسلولی بودن (Emergence of Multicellularity)
چندسلولی بودن (Multicellularity) به صورت مستقل در چندین شاخه (Lineages) تکامل یافت، از جمله:
-
حیوانات (Animals)
-
گیاهان (Plants)
-
قارچها (Fungi)
-
جلبکها (Algae)
این گذار شامل تکامل مکانیزمهای چسبندگی سلولی (Cell Adhesion)، ارتباط سلولی (Cell Communication) و تمایز سلولی (Differentiation) بود، که امکان عملکرد گروهی سلولها بهعنوان ارگانیسمهای یکپارچه (Integrated Organisms) را فراهم میکرد.
چندسلولی بودن تخصصی شدن بافتها و اندامها (Specialization of Tissues and Organs) را ممکن کرد و منجر به اندازههای بزرگتر و مورفولوژیهای پیچیدهتر (Complex Morphologies) شد. شواهد فسیلی مانند زیستبوم ایدیاکاران ۶۰۰ میلیون ساله (600-Million-Year-Old Ediacaran Biota) نشاندهنده اولین نمونههای زندگی چندسلولی با بدن نرم و شکلهای نسبتاً ساده است.
ظهور چندسلولی بودن یک نوآوری عمده (Major Innovation) بود که زمینه را برای اکوسیستمهای پیچیده در دورههای زمینشناسی بعدی فراهم کرد.
۶.۴ انفجار کامبرین (The Cambrian Explosion)
حدود ۵۴۱ میلیون سال پیش، انفجار کامبرین (Cambrian Explosion) دورهای از تنوع سریع در زندگی حیوانی (Rapid Diversification in Animal Life) بود. در بازه زمانی زمینشناسی نسبتاً کوتاه، اکثر شاخههای اصلی حیوانی (Major Animal Phyla) ظاهر شدند و طرحهای بدنی، استراتژیهای تغذیهای و نقشهای اکولوژیکی متنوع را نشان دادند.
فسیلهای این دوره، مانند نمونههای برگرفته از بورگس شیل (Burgess Shale)، تنوع مورفولوژیکی فوقالعادهای را نمایش میدهند، از جمله:
-
اولین شواهد چشمهای پیچیده (Complex Eyes)
-
اندامهای شکارچی (Predatory Appendages)
-
اسکلتهای سخت (Hardened Exoskeletons)
عوامل متعددی به انفجار کامبرین کمک کردند، از جمله:
-
افزایش دسترسی به اکسیژن (Increased Oxygen Availability)
-
تکامل ژنهای توسعهای (Developmental Genes) که طرحهای بدنی جدید را ممکن کردند
-
تعاملات اکولوژیکی مانند رابطه شکارچی-شکار (Predator-Prey Dynamics)
این رویداد نشان میدهد که فرآیندهای تکاملی میتوانند تحت شرایط محیطی و ژنتیکی مطلوب، نوآوریهای سریع ایجاد کنند.
۶.۵ سکونت خشکی (Colonization of Land)
انتقال از محیطهای آبی به خشکی (Aquatic to Terrestrial Environments) به طور مستقل در چندین شاخه رخ داد، از جمله:
-
گیاهان (Plants)
-
بندپایان (Arthropods)
-
مهرهداران (Vertebrates)
گیاهان اولیه سازگاریهایی برای جلوگیری از خشکی (Desiccation), حمایت ساختاری و تولیدمثل بدون آب ایجاد کردند، که منجر به ظهور خزهها، سرخسها و در نهایت گیاهان دانهدار (Seed Plants) شد.
بندپایان اسکلت خارجی (Exoskeleton)، سیستمهای نای (Tracheal Systems) و اندامهای مناسب برای حرکت در خشکی توسعه دادند.
مهرهداران از جمله دوزیستان اولیه (Early Amphibians)، اندامهایی برای تحمل وزن در خشکی، شش برای تنفس هوا و سازگاریهای تولیدمثلی خارج از محیط آبی پیدا کردند.
سکونت خشکی به طور چشمگیری فرصتهای اکولوژیکی را گسترش داد و منجر به شکلگیری اکوسیستمهای پیچیده خشکی شد.
۶.۶ تکامل مهرهداران (Vertebrate Evolution)
مهرهداران دستخوش یک سری گذار تکاملی عمده (Major Evolutionary Transitions) شدند، از جمله:
-
ماهیان بدون فک (Jawless Fish)
-
ماهیان فکدار (Jawed Fish)
-
چهاراندامها (Tetrapods) قابل زندگی در خشکی
تکامل فکها (Jaws) امکان استراتژیهای تغذیهای مؤثرتر و شکار را فراهم کرد، در حالی که بالهها و اندامهای جفتشده (Paired Fins and Limbs) حرکت در زیستگاههای متنوع را ممکن ساختند.
در طول صدها میلیون سال، مهرهداران به دوزیستان، خزندگان، پرندگان و پستانداران تنوع یافتند، که هر گروه سازگاریهای منحصر به فرد (Unique Adaptations) با محیطهای خود داشت.
نوآوریهای کلیدی شامل:
-
اندوترمی در پرندگان و پستانداران (Endothermy)
-
تخمهای آمنیوتیک در خزندگان (Amniotic Eggs)
-
سیستمهای حسی تخصصی (Specialized Sensory Systems)
مهرهداران را قادر ساخت تا در مجموعه وسیعی از نیچهای اکولوژیکی (Ecological Niches) جای بگیرند.
۶.۷ تکامل پستانداران و نخستیسانان (Mammalian and Primate Evolution)
پس از انقراض دایناسورهای غیرپرنده (Non-Avian Dinosaurs) حدود ۶۶ میلیون سال پیش، پستانداران یک تابش تطبیقی سریع (Rapid Adaptive Radiation) تجربه کردند و به شکلها و نقشهای اکولوژیکی متنوع تبدیل شدند.
تکامل نخستیسانان (Primate Evolution) با ویژگیهایی مانند:
-
دستهای چنگکی (Grasping Hands)
-
دید استریوسکوپیک (Stereoscopic Vision)
-
مغز بزرگتر (Enlarged Brains)
منجر به ظهور خط تکاملی انسان (Human Lineage) شد.
شواهد فسیلی یک رشته گونههای هومینین (Hominin Species) با افزایش ظرفیت جمجمه، راه رفتن دوپا و استفاده از ابزار را نشان میدهد که در نهایت به Homo sapiens مدرن میرسد.
ژنومیک تطبیقی (Comparative Genomics) این روابط را تأیید میکند و الگوهای اجدادی مشترک و زمانهای انشعاب (Shared Ancestry and Divergence Times) را در میان نخستیسانان آشکار میسازد.
تکامل انسان (Human Evolution) نمونهای از تأثیر تجمعی جهش، انتخاب و عوامل فرهنگی (Cumulative Effects of Mutation, Selection, and Cultural Influences) بر شکلدهی یک خط تکاملی در طول میلیونها سال است.
۶.۸ انقراضهای جمعی و فرصتهای تکاملی (Mass Extinctions and Evolutionary Opportunities)
در طول تاریخ زمین، رویدادهای انقراض جمعی (Mass Extinction Events) بارها مسیر حیات را تغییر دادهاند.
-
انقراض پرمین-تریاس (Permian-Triassic Extinction)
-
انقراض کرتاسه-پالئوژن (Cretaceous-Paleogene Extinction)
این رویدادها بخش زیادی از گونهها را حذف کرده و فرصتهای اکولوژیکی جدیدی برای تنوعیابی بازماندگان فراهم کردند.
این دورهها نشان میدهند که انقراض و فرصت تکاملی به هم پیوستهاند (Extinction and Evolutionary Opportunity Are Intertwined)؛ رویدادهای فاجعهبار میتوانند تابش تطبیقی (Adaptive Radiations) را تسریع کرده و گروههای کاملاً جدید ایجاد کنند.
درک انقراضهای جمعی بینشهایی درباره تابآوری و آسیبپذیری حیات (Resilience and Vulnerability of Life) ارائه میدهد، چه در گذشته و چه در زمینه فشارهای انسانساخت فعلی (Anthropogenic Pressures).
۶.۹ جمعبندی رویدادهای اصلی تکاملی (Synthesis of Major Evolutionary Events)
در مجموع، این نقاط عطف تکاملی نشاندهنده تعامل پویا بین نوآوری ژنتیکی، تغییرات محیطی و تعاملات اکولوژیکی (Dynamic Interplay Between Genetic Innovation, Environmental Change, and Ecological Interactions) است.
-
منشاء حیات: اولین سیستمهای تکثیر شونده را ایجاد کرد.
-
تکامل یوکاریوتها و چندسلولی بودن: پیچیدگی ساختاری و عملکردی را ممکن ساخت.
-
تابشهای تطبیقی، شامل انفجار کامبرین و تنوع بعد از انقراضها: ظرفیت نوآوری سریع را در شرایط مساعد نشان میدهد.
-
ظهور مهرهداران، پستانداران و انسان: تأثیر تجمعی فرآیندهای تکاملی در مقیاس زمینشناسی را نشان میدهد.
مطالعه این رویدادها الگوها و مکانیزمهای تکامل را روشن میکند و چارچوبی جامع برای درک تاریخ حیات روی زمین فراهم میآورد.
۸. نظریه تکامل مدرن و کاربردهای آن (Modern Evolutionary Theory and Applications)
نظریه تکامل مدرن (Modern Evolutionary Theory) مفاهیم کلاسیک داروینی (Darwinian Concepts) را با پیشرفتهای ژنتیک (Genetics)، زیستشناسی مولکولی (Molecular Biology)، بومشناسی (Ecology) و مدلسازی محاسباتی (Computational Modeling) تلفیق میکند تا چارچوبی جامع برای درک تنوع و پویایی زندگی (Life’s Diversity and Dynamics) فراهم آورد.
علاوه بر قدرت توضیحی (Explanatory Power)، نظریه تکامل کاربردهای عملی در پزشکی (Medicine)، کشاورزی (Agriculture)، حفاظت محیطزیست (Conservation) و بیوتکنولوژی (Biotechnology) دارد. با ترکیب نظریه، پژوهش تجربی و مدلسازی پیشبینیکننده، زیستشناسی تکاملی معاصر (Contemporary Evolutionary Biology) به سؤالات علمی بنیادی و چالشهای اجتماعی (Fundamental Scientific Questions and Societal Challenges) پاسخ میدهد.
۸.۱ تلفیق مدرن (The Modern Synthesis)
تلفیق مدرن (Modern Synthesis) که در اوایل تا اواسط قرن بیستم توسعه یافت، نظریه انتخاب طبیعی داروین (Darwin’s Theory of Natural Selection) را با ژنتیک مندلی (Mendelian Genetics) یکپارچه کرد.
پیشگامانی مانند رونالد فیشر (Ronald Fisher)، جی. بی. اس. هالدن (J. B. S. Haldane) و سوئال رایت (Sewall Wright) پایه ژنتیکی تغییرات تکاملی را Formalized) کردند و نشان دادند که جهش (Mutation)، بازترکیبی ژنتیکی (Recombination)، جریان ژنی (Gene Flow) و رانش ژنتیکی (Genetic Drift) چگونه فرکانس آللها (Allele Frequencies) را در جمعیتها تحت تأثیر قرار میدهند.
این تلفیق فرایندهای میکروتکاملی (Microevolutionary Processes) را با الگوهای ماکروتکاملی (Macroevolutionary Patterns) مشاهده شده در سوابق فسیلی (Fossil Record) سازگار کرد و چارچوبی منسجم برای درک گونهزایی (Speciation)، سازگاری (Adaptation) و تنوع فیلوژنتیکی (Phylogenetic Diversification) ارائه داد.
با پیوند ویژگیهای قابل مشاهده (Observable Traits) به مکانیزمهای ژنتیکی پایهای (Underlying Genetic Mechanisms)، تلفیق مدرن زیستشناسی تکاملی را از علمی توصیفی به علمی پیشبینیکننده و کمی (Predictive, Quantitative Discipline) تبدیل کرد.
نکته مهم: این تلفیق نشان میدهد که تکامل میتواند به طور کمی و با پیشبینی مدل شود و نه فقط توصیفی باشد.
۸.۲ تلفیق تکاملی توسعهیافته (Extended Evolutionary Synthesis – EES)
تحولات اخیر، که اغلب تلفیق تکاملی توسعهیافته (Extended Evolutionary Synthesis, EES) نامیده میشوند، دیدگاههای کلاسیک را گسترش میدهند و پدیدههایی مانند:
-
وارثت اپیژنتیکی (Epigenetic Inheritance)
-
انعطافپذیری فنوتیپی (Phenotypic Plasticity)
-
گرایشهای توسعهای (Developmental Bias)
-
ساختگاه زیستی (Niche Construction)
را در نظر میگیرند.
توضیح نکات مهم:
-
تغییرات اپیژنتیکی (Epigenetic Modifications): میتوانند بیان ژنها را بدون تغییر توالی DNA تغییر دهند و نشان میدهند که تنوع قابل ارث (Heritable Variation) میتواند فراتر از جهشهای مرسوم (Conventional Mutation) ایجاد شود.
-
انعطافپذیری فنوتیپی (Phenotypic Plasticity): به ارگانیسمها اجازه میدهد تا ویژگیها را بر اساس شرایط محیطی تغییر دهند و مسیرهای انتخاب طبیعی و تکاملی را تحت تأثیر قرار دهند.
-
ساختگاه زیستی (Niche Construction): بر این نکته تأکید دارد که ارگانیسمها محیط خود را فعالانه تغییر میدهند و به این ترتیب فشارهای انتخابی (Selection Pressures) را بر خود و گونههای دیگر شکل میدهند.
این توسعهها دیدگاه کلاسیک را تکمیل میکنند و پیچیدگی تعامل بین ژنها، توسعه، بومشناسی و رفتار (Genes, Development, Ecology, and Behavior) را نشان میدهند.
۸.۳ پزشکی تکاملی (Evolutionary Medicine)
پزشکی تکاملی (Evolutionary Medicine) اصول تکاملی را برای درک سلامت، بیماری و فیزیولوژی انسان (Health, Disease, and Human Physiology) به کار میبرد.
بسیاری از بیماریها، از جمله:
-
بیماریهای عفونی (Infectious Diseases)
-
سرطان (Cancer)
-
اختلالات متابولیک (Metabolic Disorders)
تحت تأثیر فرایندهای تکاملی (Evolutionary Processes) هستند.
نکته مهم:
-
پاتوژنها (Pathogens) تحت فشارهای انتخابی میزبان (Selective Pressures Imposed by Host Immunity) و مداخلات ضد میکروبی (Antimicrobial Interventions) تکامل مییابند، که منجر به:
-
مقاومت آنتیبیوتیکی (Antibiotic Resistance)
-
تغییر آنتیژنی ویروسها (Viral Antigenic Drift)
-
ظهور بیماریهای نوظهور (Emerging Infectious Diseases)
-
دیدگاههای تکاملی همچنین آسیبپذیریهای انسان (Human Vulnerabilities)، مانند عدم تطابق بین سازگاریهای اجدادی و محیط مدرن (Mismatch between Ancestral Adaptations and Modern Environments) را روشن میکند، که مثالهایی چون چاقی، دیابت و بیماریهای قلبی-عروقی دارد.
با درک منشأ تکاملی ویژگیهای مرتبط با سلامت و پاتوژنها (Evolutionary Origins of Health-Related Traits and Pathogens)، پزشکی تکاملی میتواند استراتژیهای پیشگیری، مداخلات درمانی و سیاستهای بهداشت عمومی (Prevention Strategies, Therapeutic Interventions, and Public Health Policies) را هدایت کند.
۸.۴ بومشناسی حفاظتی و مدیریت تکاملی (Conservation Biology and Evolutionary Management)
اصول تکاملی مرکز بومشناسی حفاظتی (Conservation Biology) هستند و مدیریت گونههای در خطر و اکوسیستمها (Endangered Species and Ecosystems) را هدایت میکنند.
عوامل حیاتی برای پایداری بلندمدت گونهها (Long-Term Persistence of Species) شامل:
-
تنوع ژنتیکی (Genetic Diversity)
-
ساختار جمعیت (Population Structure)
-
پتانسیل سازگاری (Adaptive Potential)
است.
استراتژیهای حفاظتی معمولاً از مفاهیم تکاملی بهره میبرند، از جمله:
-
حفظ جریان ژنی بین جمعیتهای پراکنده (Gene Flow)
-
مدیریت رخدادهای اختلاط ژنتیکی (Hybridization Events)
-
پیشبینی پاسخها به تغییرات اقلیمی (Responses to Climate Change)
درک تاریخ تکاملی گونهها (Evolutionary History) همچنین به اولویتبندی اقدامات حفاظتی (Conservation Prioritization) کمک میکند، از جمله شناسایی گونهها یا جمعیتهایی با شاخههای تکاملی منحصربهفرد (Unique Evolutionary Lineages) یا اهمیت سازگاری بالا (High Adaptive Significance).
نکته مهم: تلفیق تکامل در برنامهریزی حفاظتی تضمین میکند که تنوع زیستی نه تنها از نظر تعداد گونهها بلکه از نظر تنوع ژنتیکی و عملکردی (Genetic and Functional Diversity) حفظ شود.
۸.۵ کشاورزی و انتخاب مصنوعی (Agriculture and Artificial Selection)
انتخاب مصنوعی (Artificial Selection) در کشاورزی و دامپروری، کاربرد مستقیم اصول تکاملی (Direct Application of Evolutionary Principles) است.
انسانها از دیرباز ویژگیهای مطلوب در گیاهان، دامها و حیوانات اهلی (Crops, Livestock, and Domesticated Animals) را انتخاب کرده و ژنتیک آنها را طی نسلها شکل دادهاند.
برنامههای پرورشی مدرن شامل:
-
ژنتیک کمی (Quantitative Genetics)
-
نشانههای مولکولی (Molecular Markers)
-
ویرایش ژنوم (Genome Editing)
میشوند تا نتایج مطلوب مانند مقاومت به بیماری، بهینهسازی محصول و تابآوری محیطی (Disease Resistance, Yield Optimization, Environmental Resilience) را سریعتر به دست آورند.
این رویکردها نشان میدهند که درک مکانیزمهای تکاملی به انسانها اجازه میدهد تغییرات صفاتی را در جمعیتهای کنترلشده پیشبینی و هدایت کنند، مشابه فرایندهای تکاملی طبیعی (Natural Evolutionary Processes) اما با سرعت و هدفمندی بیشتر (Accelerated and Targeted Scale).
۸.۶ بیوتکنولوژی و تکامل هدایتشده (Biotechnology and Directed Evolution)
تکامل هدایتشده (Directed Evolution) روشی در بیوتکنولوژی (Biotechnology) است که انتخاب طبیعی را شبیهسازی میکند (Mimics Natural Selection) تا پروتئینها، آنزیمها و مسیرهای متابولیک (Proteins, Enzymes, and Metabolic Pathways) با عملکرد جدید یا بهبود یافته ایجاد شود.
با ایجاد جهشها (Inducing Mutations)، انتخاب ویژگیهای مطلوب (Selecting for Desired Traits) و تکرار فرایند (Iterating the Process)، دانشمندان مولکولهایی با وظایف خاص در صنایع، پزشکی یا تحقیقاتی (Industrial, Medical, or Research Contexts) میسازند.
مثالها:
-
آنزیمهای با کارایی کاتالیزوری بهبود یافته (Improved Catalytic Efficiency)
-
پروتئینهای درمانی با پایداری افزایش یافته (Enhanced Stability)
-
میکروبهای مهندسی شده برای بازسازی زیستی (Bioremediation) یا تولید سوخت زیستی (Biofuel Production)
نکته مهم: تکامل هدایتشده نشان میدهد که اصول تکامل را میتوان عمداً به کار گرفت و قدرت پیشبینی مکانیزمهای جهش، انتخاب و سازگاری (Mutation, Selection, and Adaptation) را به نمایش میگذارد.
۸.۷ محاسبات و مدلسازی تکاملی (Evolutionary Computation and Modeling)
رویکردهای محاسباتی الهامگرفته از تکامل، مانند:
-
الگوریتمهای تکاملی (Evolutionary Algorithms)
-
الگوریتمهای ژنتیکی (Genetic Algorithms)
-
شبیهسازی حیات مصنوعی (Artificial Life Simulations)
اصل تکامل را به حل مسائل در علوم کامپیوتر، مهندسی و بهینهسازی (Problem-Solving in Computer Science, Engineering, and Optimization) گسترش میدهند.
این الگوریتمها جهش، بازترکیبی، انتخاب و ارزیابی سازگاری (Fitness Evaluation) را شبیهسازی میکنند تا راهحلهای مسائل پیچیده را بهبود دهند (Iteratively Improve Solutions).
کاربردها:
-
بهینهسازی طراحی مهندسی (Engineering Designs)
-
آموزش مدلهای یادگیری ماشین (Machine Learning Models)
-
شبیهسازی سناریوهای بومشناختی یا تکاملی (Ecological or Evolutionary Scenarios)
نکته مهم: محاسبات تکاملی نشان میدهد که اصول تکامل تنها به زیستشناسی محدود نیستند و فرایندهای مبتنی بر انتخاب (Selection-Driven Processes) قابلیت کاربرد گسترده دارند.
۸.۸ درک تکامل معاصر (Understanding Contemporary Evolution)
زیستشناسی تکاملی مدرن همچنین تغییرات تکاملی سریع و جاری (Ongoing and Rapid Evolutionary Changes) در محیطهای طبیعی و انسانی (Natural and Anthropogenic Contexts) را بررسی میکند.
فشارهای انتخابی جدید شامل:
-
تغییرات اقلیمی (Climate Change)
-
تکهتکه شدن زیستگاهها (Habitat Fragmentation)
-
آلودگی (Pollution)
-
شهرنشینی (Urbanization)
نتایج این فشارها شامل تغییرات قابل اندازهگیری در مورفولوژی، رفتار و ژنتیک (Morphology, Behavior, and Genetics) طی زمانهای کوتاه است.
نکته مهم: مطالعات تکامل معاصر بینشهایی درباره ظرفیت سازگاری (Adaptive Capacity)، تابآوری (Resilience) و پیامدهای بالقوه تغییر محیطی (Potential Consequences of Environmental Change) ارائه میدهند و اهمیت نظریه تکامل در مواجهه با چالشهای جهانی فوری (Urgent Global Challenges) را برجسته میکنند.
۸.۹ تلفیق کاربردهای تکاملی مدرن (Synthesis of Modern Evolutionary Applications)
تلفیق مدرن (Modern Synthesis)، چارچوبهای تکاملی توسعهیافته (Extended Evolutionary Frameworks) و پژوهشهای کاربردی تکاملی (Applied Evolutionary Research) به طور جمعی اهمیت پایدار نظریه تکامل (Enduring Relevance of Evolutionary Theory) را نشان میدهند.
با ترکیب اصول کلاسیک با:
-
ژنتیک مولکولی (Molecular Genetics)
-
ابزارهای محاسباتی (Computational Tools)
-
دیدگاههای بومشناختی (Ecological Perspectives)
زیستشناسی تکاملی معاصر قدرت توضیحی و کاربرد عملی (Explanatory Power and Practical Utility) فراهم میکند.
نمونه کاربردها:
-
پزشکی (Medicine)
-
حفاظت محیط زیست (Conservation)
-
کشاورزی (Agriculture)
-
بیوتکنولوژی (Biotechnology)
-
مدلسازی محاسباتی (Computational Modeling)
۹. نتیجهگیری و چشماندازهای آینده (Conclusion and Future Directions)
زیستشناسی تکاملی (Evolutionary Biology) به یکی از جامعترین و توضیحدهندهترین چارچوبها در علوم زیستی (Integrative and Explanatory Frameworks in Life Sciences) تبدیل شده است و بینشهای عمیقی درباره منشاء، تنوع و سازگاری زندگی روی زمین (Origin, Diversification, and Adaptation of Life on Earth) ارائه میدهد.
از مشاهدات اولیه داروین در مورد انتخاب طبیعی (Darwin’s Observations of Natural Selection) تا رویکردهای مدرن ژنومیک (Genomic) و محاسباتی (Computational Approaches)، مطالعه تکامل فرایندهای مولکولی (Molecular Processes)، ویژگیهای موجودات (Organismal Traits)، تعاملات بومشناختی (Ecological Interactions) و الگوهای تاریخی (Historical Patterns) را در مقیاسهای مختلف به هم پیوند میدهد.
این تلفیق (Synthesis) نه تنها پدیدههای زیستی گذشته و حال (Past and Present Biological Phenomena) را توضیح میدهد، بلکه ابزارهای پیشبینیکنندهای (Predictive Tools) برای پاسخ به چالشهای کنونی و آینده در پزشکی، حفاظت، کشاورزی و بیوتکنولوژی فراهم میکند.
۹.۱ درک جامع زندگی (Integrative Understanding of Life)
قدرت نظریه تکامل (Evolutionary Theory) در توانایی آن برای ادغام شواهد متنوع در یک چارچوب منسجم (Integrate Diverse Lines of Evidence into a Coherent Framework) نهفته است.
شواهد شامل:
-
سوابق فسیلی (Fossil Records)
-
آناتومی تطبیقی (Comparative Anatomy)
-
جنینشناسی (Embryology)
-
دادههای مولکولی (Molecular Data)
همگی با هم نشان میدهند که:
-
تبار مشترک (Common Ancestry)
-
تغییرات سازگارانه (Adaptive Change)
-
الگوهای منشعب گونهزایی (Branching Patterns of Speciation)
وجود دارد.
مطالعات مشاهداتی و تجربی (Observational and Experimental Studies) در جمعیتهای طبیعی فرایندهای تکاملی را در زمان واقعی نشان میدهند (Reveal Evolutionary Processes in Real Time)، در حالی که تحلیلهای ژنومی و محاسباتی (Genomic and Computational Analyses) بینش مکانیکی دقیق درباره پایه ژنتیکی تنوع، سازگاری و مسیرهای تکاملی (Genetic Basis of Variation, Adaptation, and Evolutionary Trajectories) ارائه میکنند.
با ترکیب این رویکردها، زیستشناسی تکاملی درکی جامع از زندگی ارائه میدهد که از سطح مولکولی تا سطح بومشناختی گسترده است (Comprehensive Understanding of Life Spanning Molecular to Ecological Scale).
نکته مهم: توانایی تلفیق شواهد از سطوح مختلف، ویژگی برجسته و قدرت توضیحی اصلی نظریه تکامل است.
۹.۲ تکامل به عنوان یک فرایند پویا (Evolution as a Dynamic Process)
تکامل تنها یک فرایند تاریخی نیست، بلکه نیروی جاری و پویا (Ongoing and Dynamic Force) است که تنوع و سازگاری موجودات (Diversity and Adaptability of Organisms) را شکل میدهد.
تحقیقات تکاملی معاصر نشان میدهد که تغییرات محیطی (Environmental Change)، فعالیتهای انسانی (Human Activity) و تأثیرات فرهنگی (Cultural Influences) چگونه واکنشهای تکاملی سریع (Rapid Evolutionary Responses) را شکل میدهند.
نمونهها شامل:
-
ظهور باکتریهای مقاوم به آنتیبیوتیک (Antibiotic-Resistant Bacteria)
-
سازگاریهای فنوتیپی در حیات وحش شهری (Phenotypic Adaptations in Urban Wildlife)
این مثالها نشان میدهند که تکامل به طور مداوم و تعاملی عمل میکند (Operates Continuously and Interactively) و هم تابآوری (Resilience) و هم آسیبپذیری (Vulnerability) سیستمهای زنده را نمایان میسازد.
نکته مهم: شناخت تکامل به عنوان یک فرایند پویا اهمیت آن را در مواجهه با چالشهای جهانی در حوزه سلامت، پایداری و مدیریت تنوع زیستی (Global Challenges in Health, Sustainability, and Biodiversity Management) تأکید میکند.
۹.۳ کاربردها در علم و جامعه (Applications in Science and Society)
اصول تکامل کاربردهایی فراتر از پژوهش دانشگاهی دارند:
-
پزشکی (Medicine):
-
راهنمایی استراتژیها برای مبارزه با بیماریهای عفونی (Combating Infectious Diseases)
-
درک اختلالات ژنتیکی (Genetic Disorders)
-
توسعه درمانهای نوین (Novel Therapeutics)
-
-
زیستشناسی حفاظتی (Conservation Biology):
-
درک تاریخ تکاملی (Evolutionary History) و پتانسیل سازگاری (Adaptive Potential)
-
حفظ تنوع ژنتیکی و تابآوری اکوسیستم (Genetic Diversity and Ecosystem Resilience)
-
-
کشاورزی و بیوتکنولوژی (Agriculture and Biotechnology):
-
بهبود محصولات کشاورزی (Crop Yield)
-
مهندسی پروتئینها (Proteins)
-
بهینهسازی سیستمهای میکروبی (Microbial Systems)
-
-
محاسبات و هوش مصنوعی (Computational and Artificial Intelligence Approaches):
-
استفاده از مفاهیم تکامل برای حل مسائل پیچیده (Problem-Solving)
-
نکته مهم: کاربرد عملی نظریه تکامل نشان میدهد که این چارچوب علمی برای حل مشکلات واقعی جهان (Solving Real-World Problems) حیاتی است.
۹.۴ چالشهای جاری و مرزهای پژوهش (Ongoing Challenges and Research Frontiers)
با وجود پیشرفتهای چشمگیر، سؤالات زیادی در زیستشناسی تکاملی باز ماندهاند.
-
مکانیزمهای تحولات بزرگ (Major Transitions) مانند:
-
منشأ زندگی (Origin of Life)
-
تکامل چندسلولی شدن (Evolution of Multicellularity)
-
ظهور رفتارهای پیچیده (Emergence of Complex Behaviors)
-
همچنان در حال تحقیق فعال هستند.
-
ادغام دادههای ژنومی، اپیژنتیکی، بومشناختی و فرهنگی (Integrating Genomic, Epigenetic, Ecological, and Cultural Data)، هم چالشهای روششناختی و مفهومی (Methodological and Conceptual Challenges) ایجاد میکند.
-
پیشبینی واکنشهای تکاملی به تغییرات سریع محیطی (Predicting Evolutionary Responses to Rapid Environmental Change) مانند:
-
تغییرات اقلیمی (Climate Change)
-
از دست رفتن زیستگاه (Habitat Loss)
-
فشارهای انتخابی ناشی از فعالیتهای انسانی (Human-Induced Selection Pressures)
-
نیازمند مدلهای پیچیده با در نظر گرفتن عوامل متقابل متعدد (Sophisticated Models Accounting for Multiple Interacting Factors) است.
نکته مهم: مقابله با این چالشها نیازمند همکاری میانرشتهای، رویکردهای آزمایشی نوآورانه و مدلسازی پیشرفته محاسباتی (Interdisciplinary Collaboration, Innovative Experimental Approaches, Advanced Computational Modeling) است.
۹.۵ آینده زیستشناسی تکاملی (The Future of Evolutionary Biology)
آینده زیستشناسی تکاملی در درک عمیقتر تعامل بین ژنها، محیط و رفتار (Interplay Between Genes, Environment, and Behavior) و تبدیل بینشهای نظری به راهحلهای عملی (Translating Theoretical Insights into Practical Solutions) نهفته است.
فناوریهای نوظهور (Emerging Technologies):
-
ژنومیک پرسرعت (High-Throughput Genomics)
-
توالییابی تکسلولی (Single-Cell Sequencing)
-
مطالعات عملکردی مبتنی بر CRISPR (CRISPR-Based Functional Studies)
-
پایش بومشناختی (Ecological Monitoring)
این فناوریها فرصتهای بیسابقهای برای مطالعه تکامل در سطوح مولکولی، موجودات و اکوسیستمها (Molecular, Organismal, and Ecosystem Levels) فراهم میکنند.
تلفیق دانش تکاملی با پزشکی، حفاظت، کشاورزی و سیستمهای مصنوعی (Artificial Systems) همچنان درک علمی و سودمندی اجتماعی (Scientific Understanding and Societal Benefit) را گسترش میدهد.
نکته مهم: با اتصال الگوهای تاریخی به مدلهای پیشبینیکننده (Linking Historical Patterns with Predictive Models)، زیستشناسی تکاملی مرکز توجه برای مقابله با چالشهای زیستی قرن ۲۱ باقی خواهد ماند.
۹.۶ تلفیق (Synthesis)
در پایان، زیستشناسی تکاملی چارچوبی متحدکننده برای درک تنوع و پیچیدگی زندگی (Unifying Framework for Understanding Diversity and Complexity of Life) ارائه میدهد.
از منشأ مولکولی تنوع (Molecular Origins of Variation) تا ظهور موجودات و اکوسیستمهای پیچیده (Emergence of Complex Organisms and Ecosystems)، فرایندهای تکاملی (Evolutionary Processes) هم تاریخ زندگی و هم وضعیت کنونی آن را شکل میدهند (Shape Both History and Present State of Life).
تلفیق دیدگاههای کلاسیک و مدرن (Classical and Modern Perspectives) همراه با کاربردهای عملی (Practical Applications) اهمیت پایدار و قدرت تحولآفرین نظریه تکامل (Enduring Relevance and Transformative Power of Evolutionary Theory) را تأکید میکند.
با پیشرفت پژوهش و فناوری، زیستشناسی تکاملی همچنان مکانیزمهای شکلدهنده تنوع زندگی را روشن و تلاشها برای حفاظت و بهبود جهان زنده را هدایت خواهد کرد (Illuminate Mechanisms Driving Life’s Diversity and Guide Efforts to Steward and Enhance the Living World).


